

Centrosome movements in vivo correlate with specific neurite formation downstream of LIM homeodomain transcription factor activity
- PMID: 22899847
- PMCID: PMC3436112
- DOI: 10.1242/dev.081513
Centrosome movements in vivo correlate with specific neurite formation downstream of LIM homeodomain transcription factor activity
Abstract
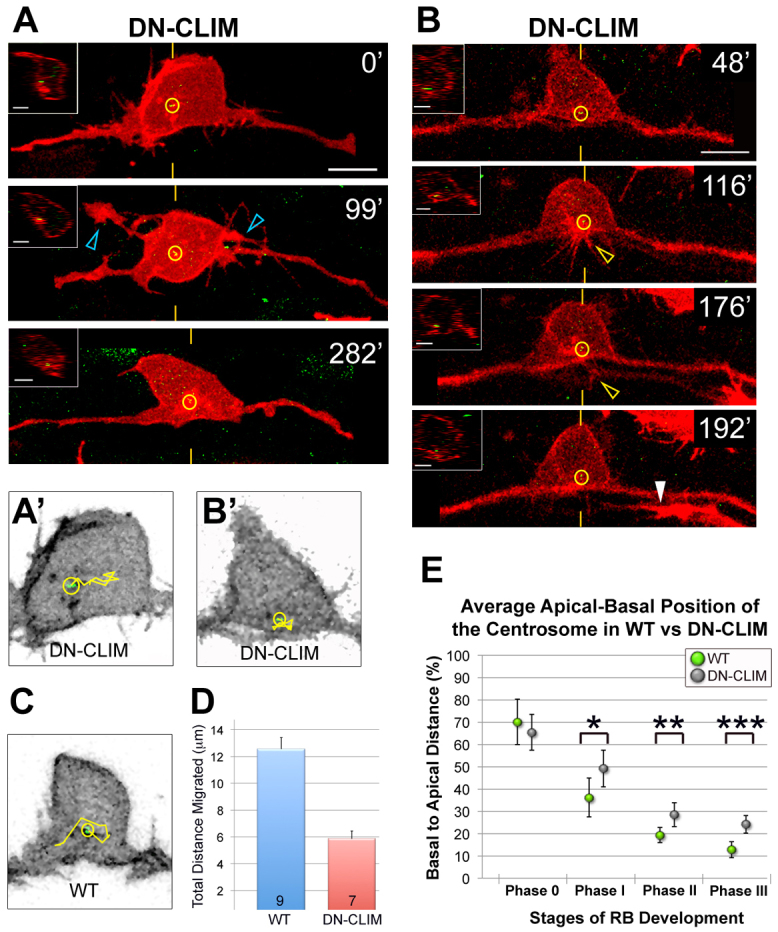
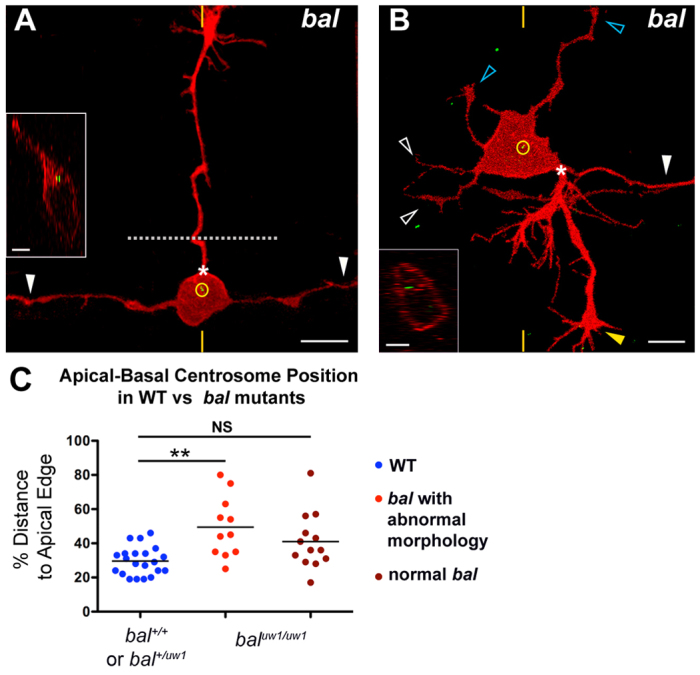
Neurons must develop complex structure to form proper connections in the nervous system. The initiation of axons in defined locations on the cell body and their extension to synaptic targets are critical steps in neuronal morphogenesis, yet the mechanisms controlling axon formation in vivo are poorly understood. The centrosome has been implicated in multiple aspects of neuronal morphogenesis; however, its function in axon development is under debate. Conflicting results from studies of centrosome function in axonogenesis suggest that its role is context dependent and underscore the importance of studying centrosome function as neurons develop in their natural environment. Using live imaging of zebrafish Rohon-Beard (RB) sensory neurons in vivo, we discovered a spatiotemporal relationship between centrosome position and the formation of RB peripheral, but not central, axons. We tested centrosome function by laser ablation and found that centrosome disruption inhibited peripheral axon outgrowth. In addition, we show that centrosome position and motility are regulated by LIM homeodomain transcription factor activity, which is specifically required for the development of RB peripheral axons. Furthermore, we show a correlation between centrosome mislocalization and ectopic axon formation in bashful (laminin alpha 1) mutants. Thus, both intrinsic transcription factor activity and extracellular cues can influence centrosome position and axon formation in vivo. This study presents the first positive association between the centrosome and axon formation in vivo and suggests that the centrosome is important for differential neurite formation in neurons with complex axonal morphologies.
Figures
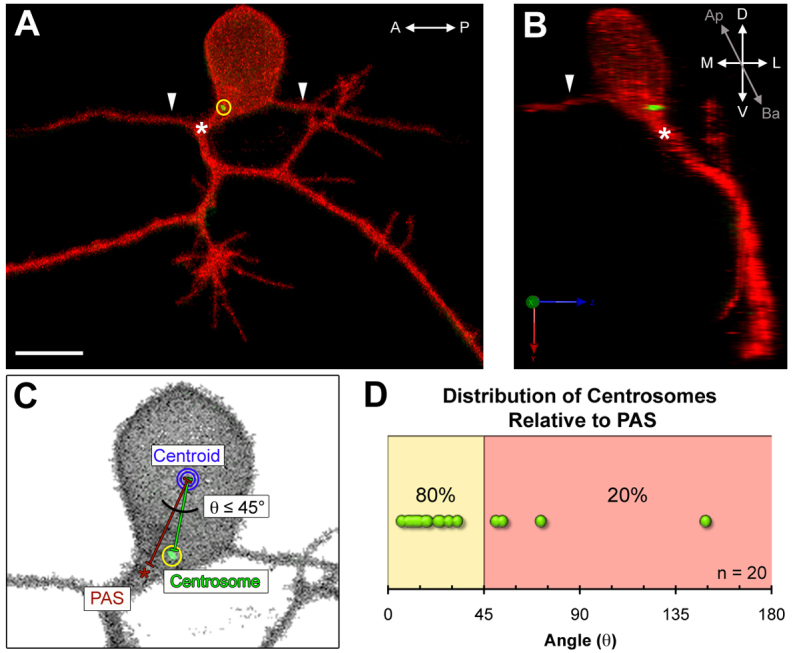
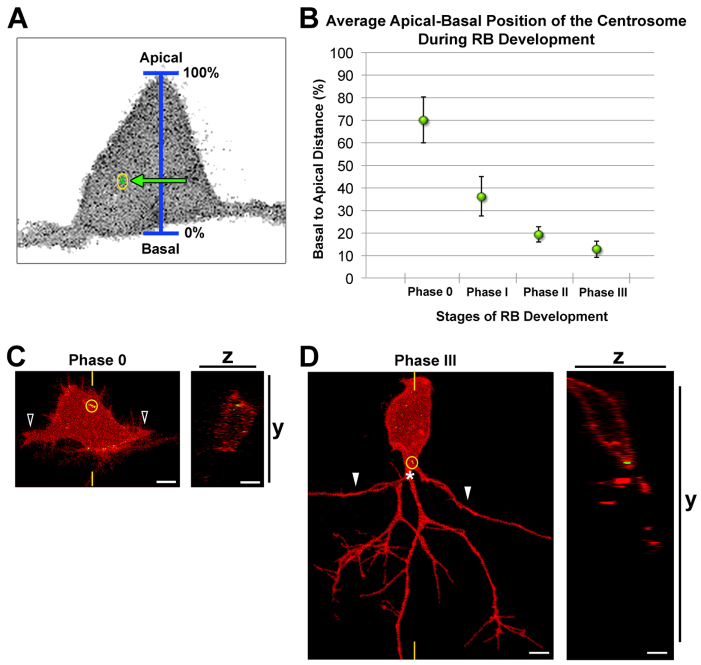
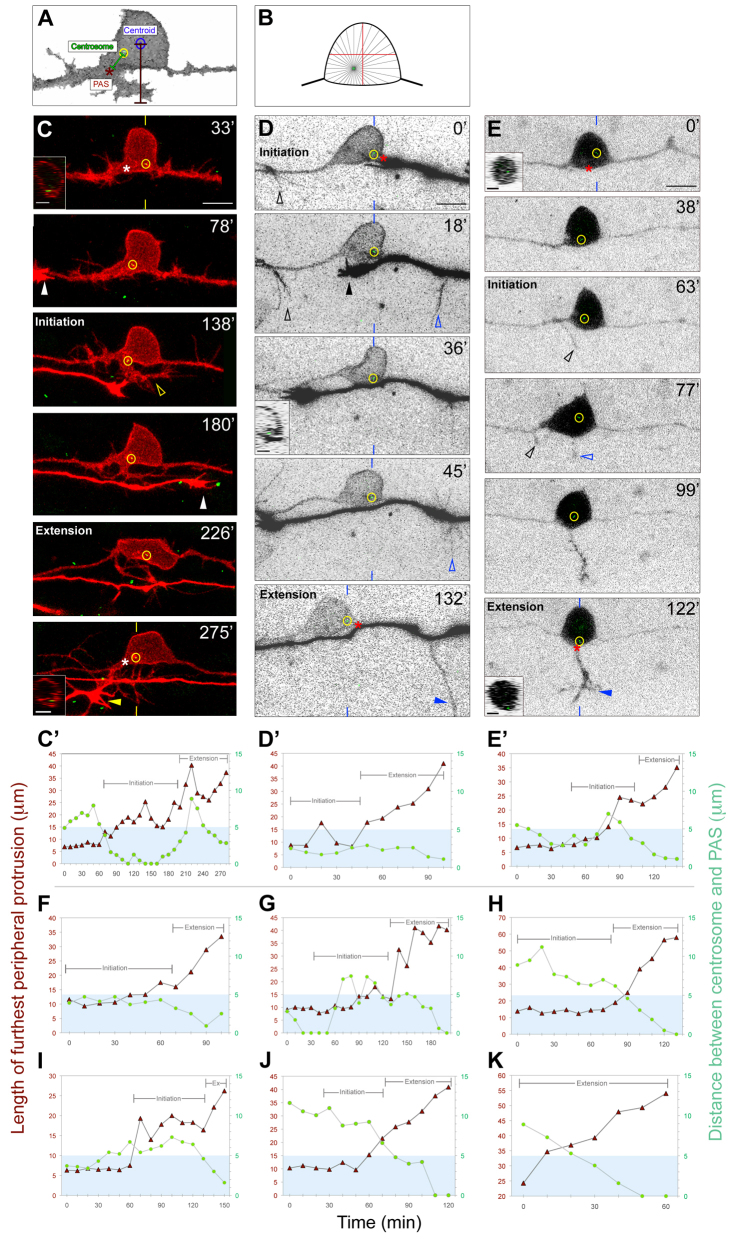
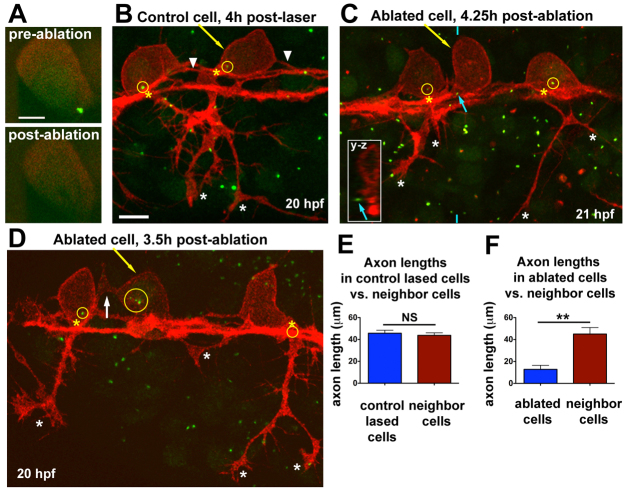
References
-
- Arimura N., Kaibuchi K. (2007). Neuronal polarity: from extracellular signals to intracellular mechanisms. Nat. Rev. Neurosci. 8, 194–205 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
