

Deciphering the complex leaf transcriptome of the allotetraploid species Nicotiana tabacum: a phylogenomic perspective
- PMID: 22900718
- PMCID: PMC3582432
- DOI: 10.1186/1471-2164-13-406
Deciphering the complex leaf transcriptome of the allotetraploid species Nicotiana tabacum: a phylogenomic perspective
Abstract
Background: Polyploidization is an important mechanism in plant evolution. By analyzing the leaf transcriptomes taken from the allotetraploid Nicotiana tabacum (tobacco) and parental genome donors, N. sylvesteris (S-Genome) and N. tomentosiformis (T-Genome), a phylogenomic approach was taken to map the fate of homeologous gene pairs in this plant.
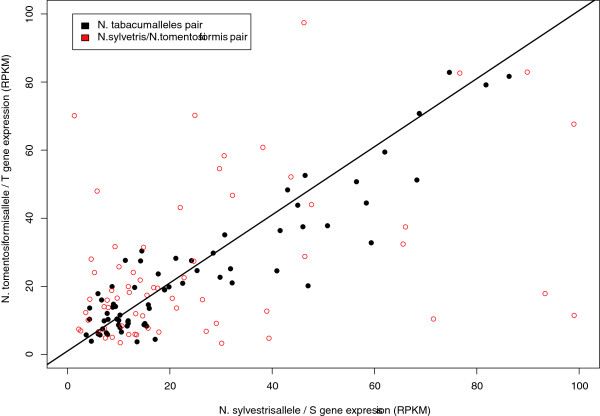
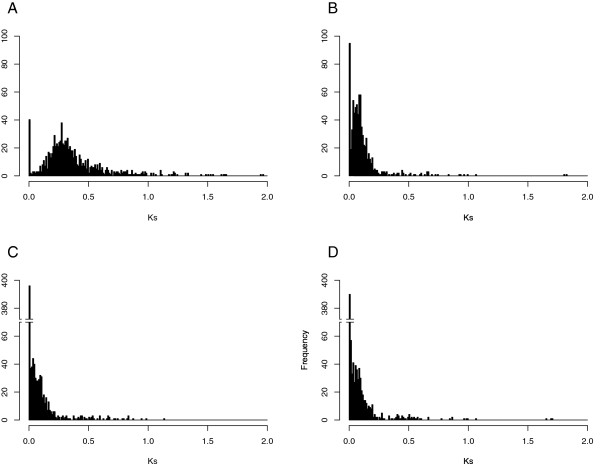
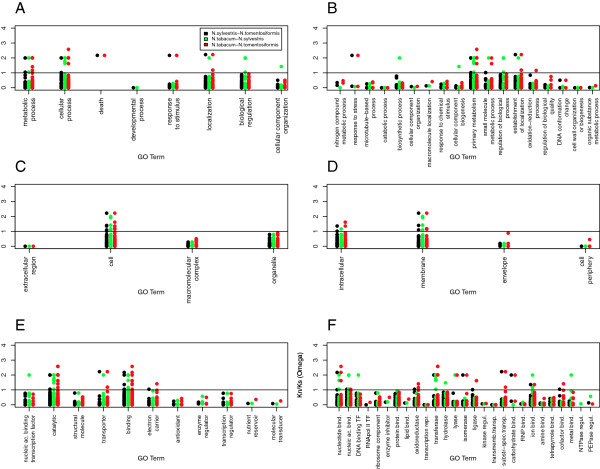
Results: A comparison between the genes present in the leaf transcriptomes of N. tabacum and modern day representatives of its progenitor species demonstrated that only 33% of assembled transcripts could be distinguished based on their sequences. A large majority of the genes (83.6% of the non parent distinguishable and 87.2% of the phylogenetic topology analyzed clusters) expressed above background level (more than 5 reads) showed similar overall expression levels. Homeologous sequences could be identified for 968 gene clusters, and 90% (6% of all genes) of the set maintained expression of only one of the tobacco homeologs. When both homeologs were expressed, only 15% (0.5% of the total) showed evidence of differential expression, providing limited evidence of subfunctionalization. Comparing the rate of synonymous nucleotide substitution (Ks) and non-synonymous nucleotide substitution (Kn) provided limited evidence for positive selection during the evolution of tobacco since the polyploidization event took place.
Conclusions: Polyploidization is a powerful mechanism for plant speciation that can occur during one generation; however millions of generations may be necessary for duplicate genes to acquire a new function. Analysis of the tobacco leaf transcriptome reveals that polyploidization, even in a young tetraploid such as tobacco, can lead to complex changes in gene expression. Gene loss and gene silencing, or subfunctionalization may explain why both homeologs are not expressed by the associated genes. With Whole Genome Duplication (WGD) events, polyploid genomes usually maintain a high percentage of gene duplicates. The data provided little evidence of preferential maintenance of gene expression from either the T- or S-genome. Additionally there was little evidence of neofunctionalization in Nicotiana tabacum suggesting it occurs at a low frequency in young polyploidy.
Figures

References
-
- Jiao Y, Wickett NJ, Ayyampalayam S, Chanderbali AS, Landherr L, Ralph PE, Tomsho LP, Hu Y, Liang H, Soltis PS, Soltis DE, Clifton SW, Schlarbaum SE, Schuster SC, Ma H, Leebens-Mack J, de Pamphilis CW. Ancestral polyploidy in seed plants and angiosperms. Nature. 2011;473(7345):97–100. doi: 10.1038/nature09916. - DOI - PubMed
-
- Jaillon O, Aury JM, Noel B, Policriti A, Clepet C, Casagrande A, Choisne N, Aubourg S, Vitulo N, Jubin C, Vezzi A, Legeai F, Hugueney P, Dasilva C, Horner D, Mica E, Jublot D, Poulain J, Bruyere C, Billault A, Segurens B, Gouyvenoux M, Ugarte E, Cattonaro F, Anthouard V, Vico V, Del Fabbro C, Alaux M, Di Gaspero G, Dumas V. French-Italian Public Consortium for Grapevine Genome Characterization et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature. 2007;449(7161):463–467. doi: 10.1038/nature06148. - DOI - PubMed
