

Disulfide bond formation in the bacterial periplasm: major achievements and challenges ahead
- PMID: 22901060
- PMCID: PMC3676657
- DOI: 10.1089/ars.2012.4864
Disulfide bond formation in the bacterial periplasm: major achievements and challenges ahead
Abstract
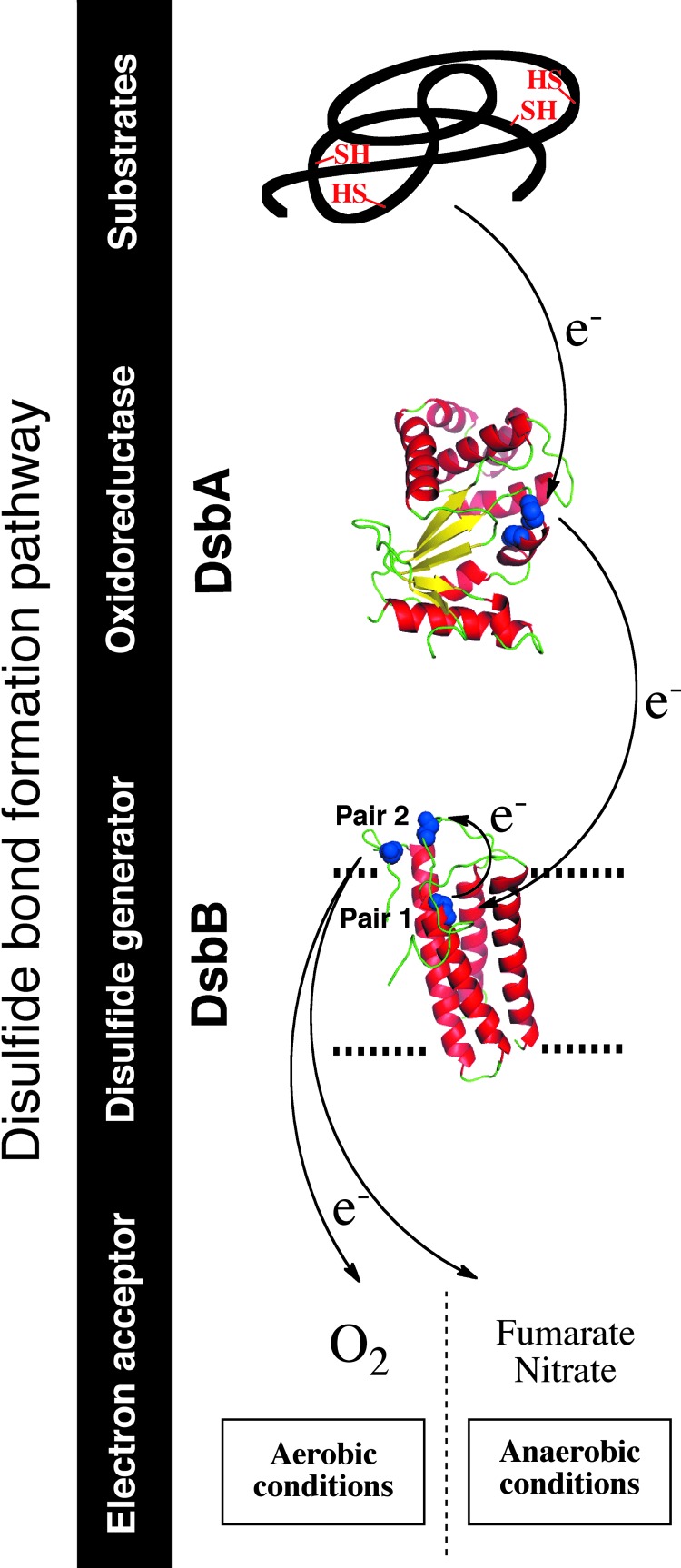
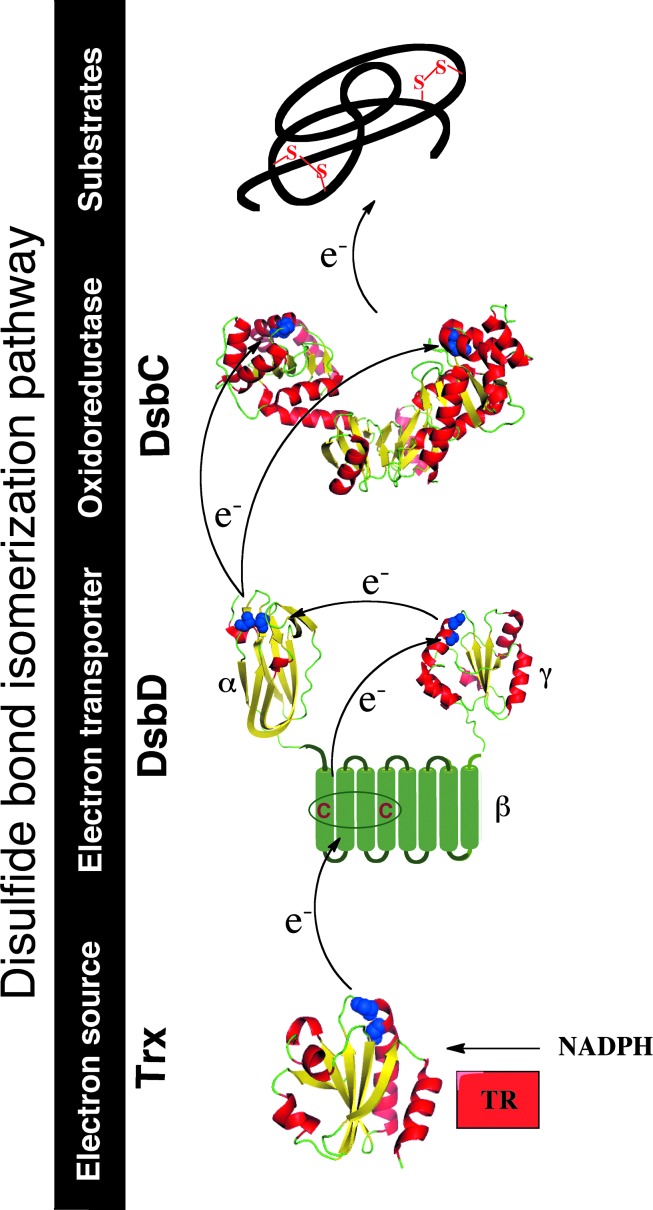
Significance: The discovery of the oxidoreductase disulfide bond protein A (DsbA) in 1991 opened the way to the unraveling of the pathways of disulfide bond formation in the periplasm of Escherichia coli and other Gram-negative bacteria. Correct oxidative protein folding in the E. coli envelope depends on both the DsbA/DsbB pathway, which catalyzes disulfide bond formation, and the DsbC/DsbD pathway, which catalyzes disulfide bond isomerization.
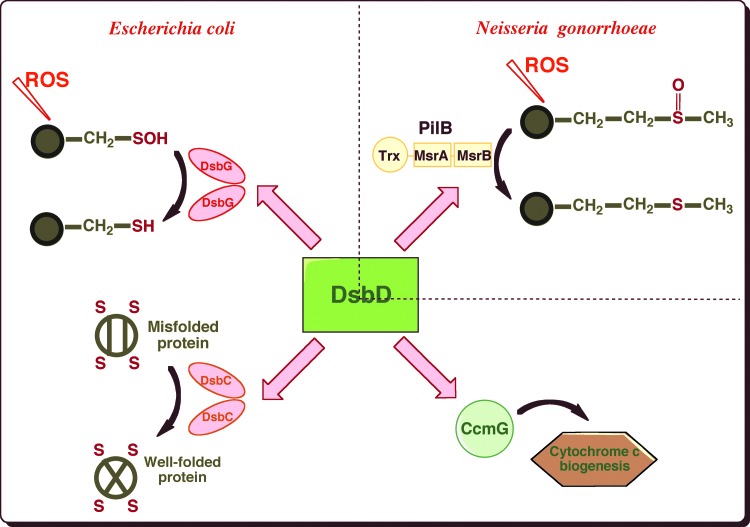
Recent advances: Recent data have revealed an unsuspected link between the oxidative protein-folding pathways and the defense mechanisms against oxidative stress. Moreover, bacterial disulfide-bond-forming systems that differ from those at play in E. coli have been discovered.
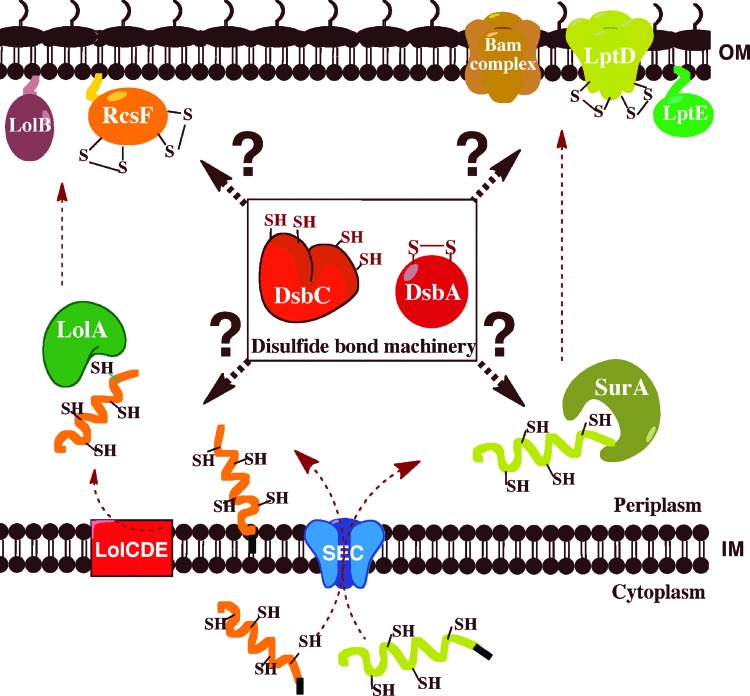
Critical issues: In this review, we discuss fundamental questions that remain unsolved, such as what is the mechanism employed by DsbD to catalyze the transfer of reducing equivalents across the membrane and how do the oxidative protein-folding catalysts DsbA and DsbC cooperate with the periplasmic chaperones in the folding of secreted proteins.
Future directions: Understanding the mechanism of DsbD will require solving the structure of the membranous domain of this protein. Another challenge of the coming years will be to put the knowledge of the disulfide formation machineries into the global cellular context to unravel the interplay between protein-folding catalysts and chaperones. Also, a thorough characterization of the disulfide bond formation machineries at work in pathogenic bacteria is necessary to design antimicrobial drugs targeting the folding pathway of virulence factors stabilized by disulfide bonds.
Figures
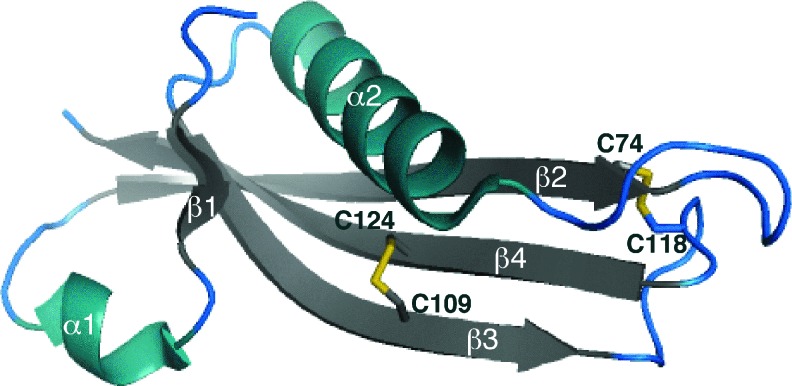
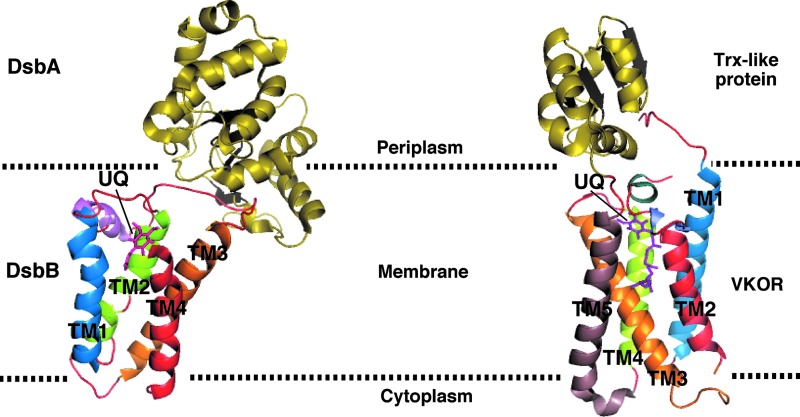
References
-
- Bader M. Muse W. Ballou DP. Gassner C. Bardwell JC. Oxidative protein folding is driven by the electron transport system. Cell. 1999;98:217–227. - PubMed
-
- Bardwell JC. McGovern K. Beckwith J. Identification of a protein required for disulfide bond formation in vivo. Cell. 1991;67:581–589. - PubMed
-
- Brot N. Collet JF. Johnson LC. Jonsson TJ. Weissbach H. Lowther WT. The thioredoxin domain of Neisseria gonorrhoeae PilB can use electrons from DsbD to reduce downstream methionine sulfoxide reductases. J Biol Chem. 2006;281:32668–32675. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
