

BIRC7-E2 ubiquitin conjugate structure reveals the mechanism of ubiquitin transfer by a RING dimer
- PMID: 22902369
- PMCID: PMC3880866
- DOI: 10.1038/nsmb.2379
BIRC7-E2 ubiquitin conjugate structure reveals the mechanism of ubiquitin transfer by a RING dimer
Abstract
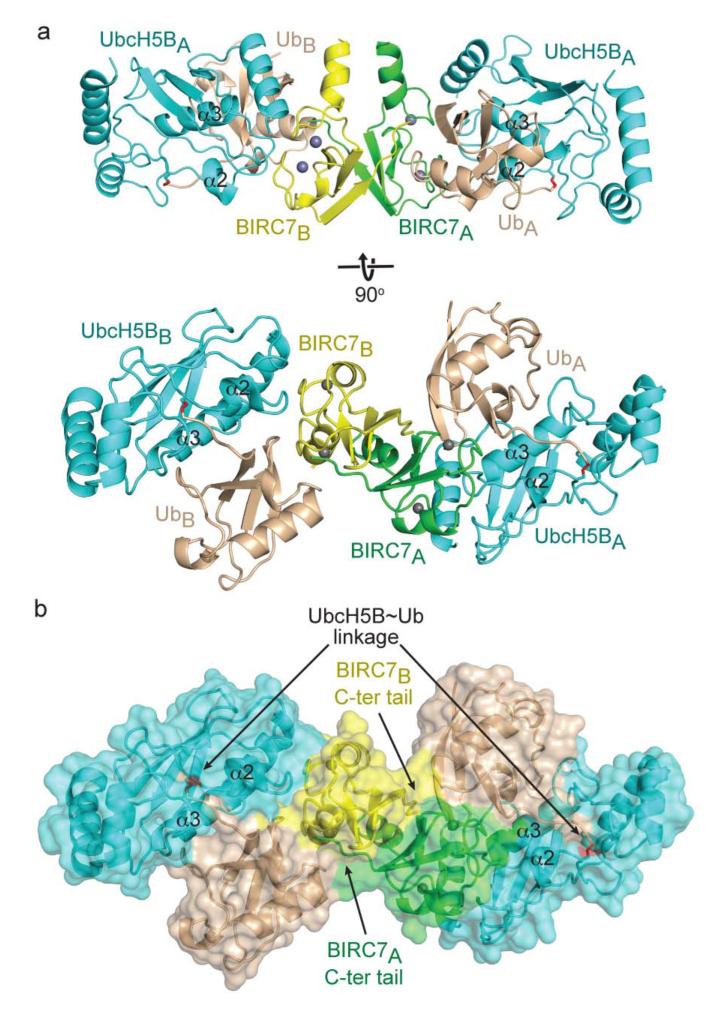
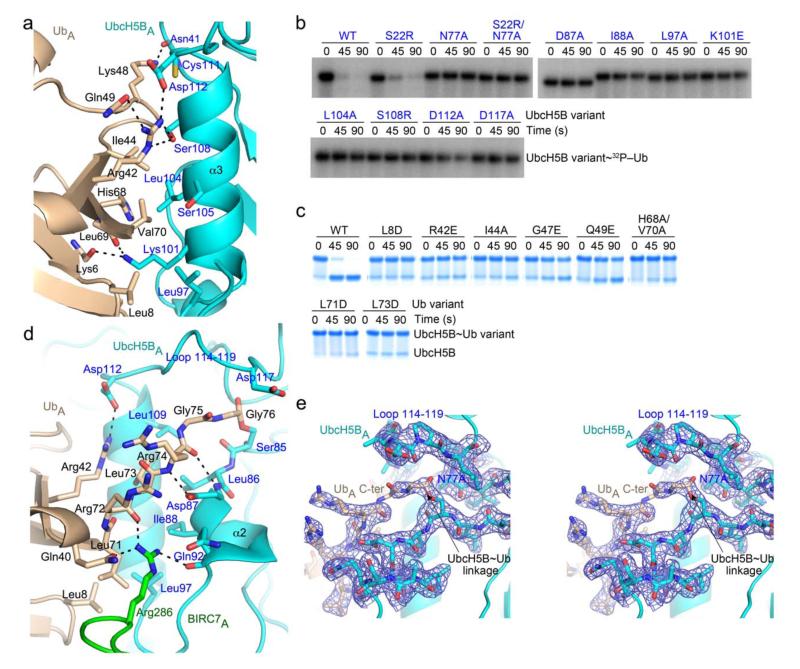
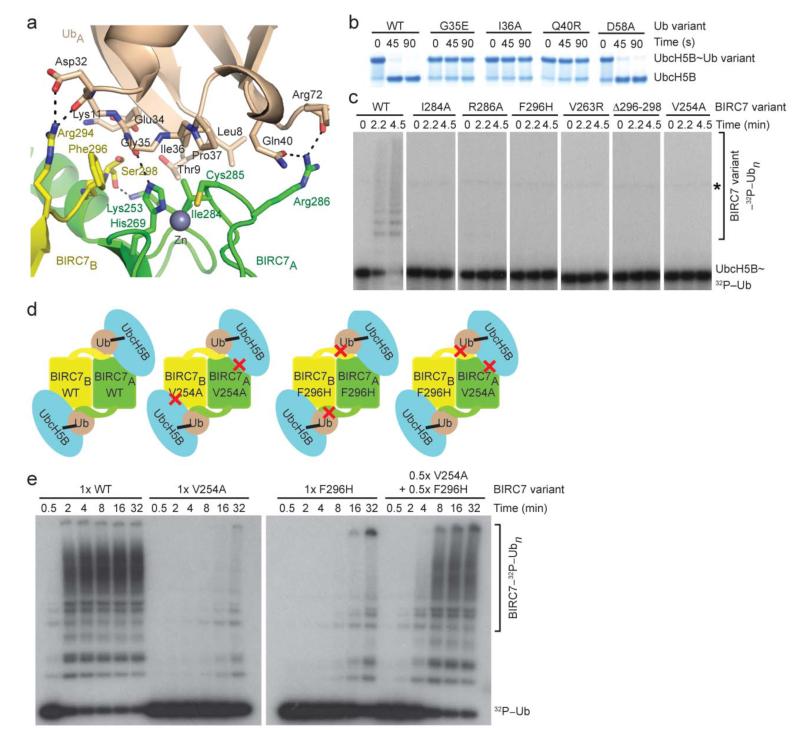
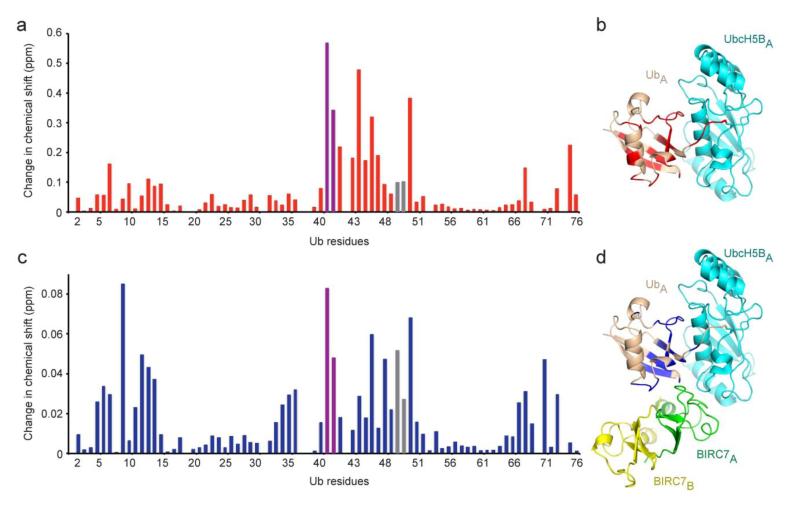
Certain RING ubiquitin ligases (E3s) dimerize to facilitate ubiquitin (Ub) transfer from ubiquitin-conjugating enzyme (E2) to substrate, but structural evidence on how this process promotes Ub transfer is lacking. Here we report the structure of the human dimeric RING domain from BIRC7 in complex with the E2 UbcH5B covalently linked to Ub (UbcH5B∼Ub). The structure reveals extensive noncovalent donor Ub interactions with UbcH5B and both subunits of the RING domain dimer that stabilize the globular body and C-terminal tail of Ub. Mutations that disrupt these noncovalent interactions or RING dimerization reduce UbcH5B∼Ub binding affinity and ubiquitination activity. Moreover, NMR analyses demonstrate that BIRC7 binding to UbcH5B∼Ub induces peak-shift perturbations in the donor Ub consistent with the crystallographically-observed Ub interactions. Our results provide structural insights into how dimeric RING E3s recruit E2∼Ub and optimize the donor Ub configuration for transfer.
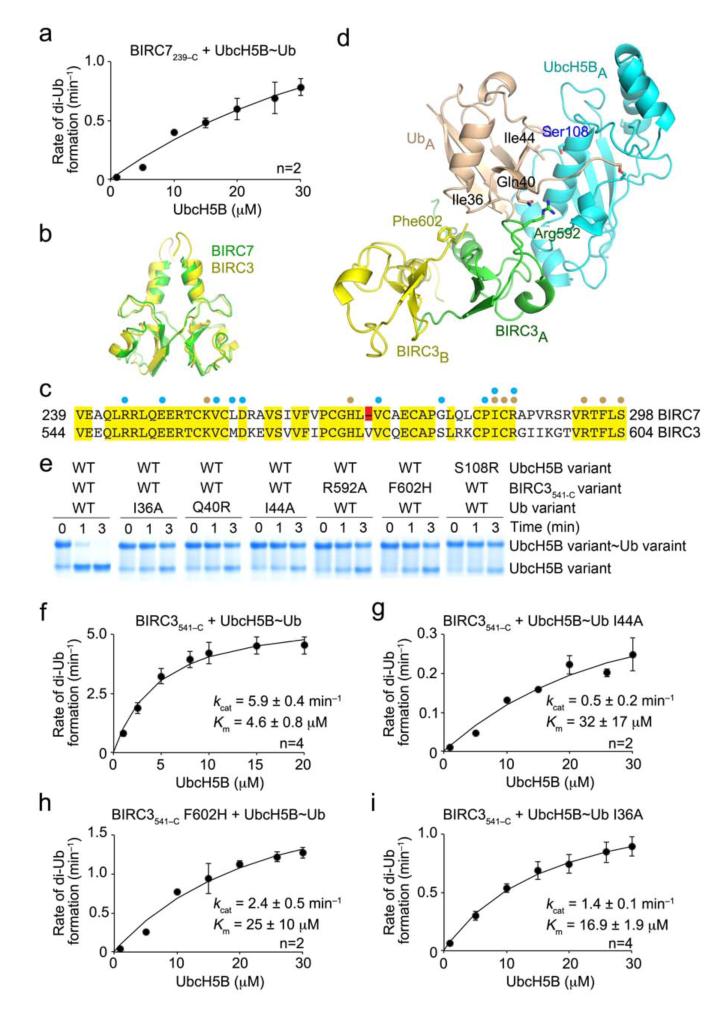
Figures
References
-
- Hershko A, Ciechanover A. The ubiquitin system. Annu Rev Biochem. 1998;67:425–79. - PubMed
-
- Pickart CM, Eddins MJ. Ubiquitin: structures, functions, mechanisms. Biochim Biophys Acta. 2004;1695:55–72. - PubMed
-
- Dye BT, Schulman BA. Structural mechanisms underlying posttranslational modification by ubiquitin-like proteins. Annu Rev Biophys Biomol Struct. 2007;36:131–50. - PubMed
-
- Deshaies RJ, Joazeiro CA. RING domain E3 ubiquitin ligases. Annu Rev Biochem. 2009;78:399–434. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
