

Rapid induction of inflammatory lipid mediators by the inflammasome in vivo
- PMID: 22902502
- PMCID: PMC3465483
- DOI: 10.1038/nature11351
Rapid induction of inflammatory lipid mediators by the inflammasome in vivo
Abstract
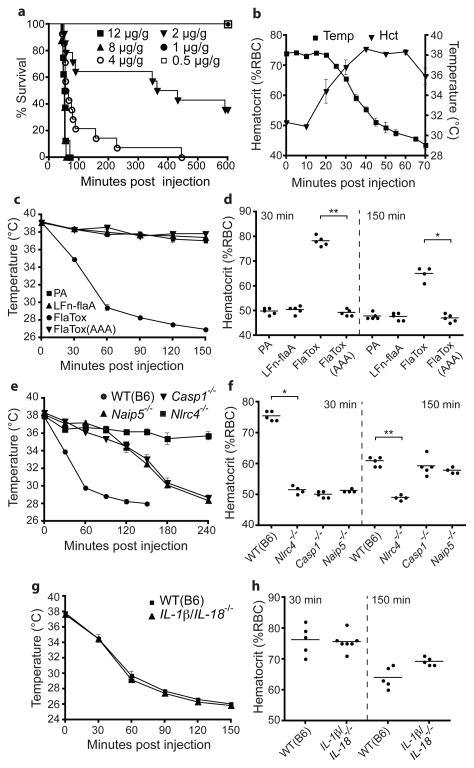
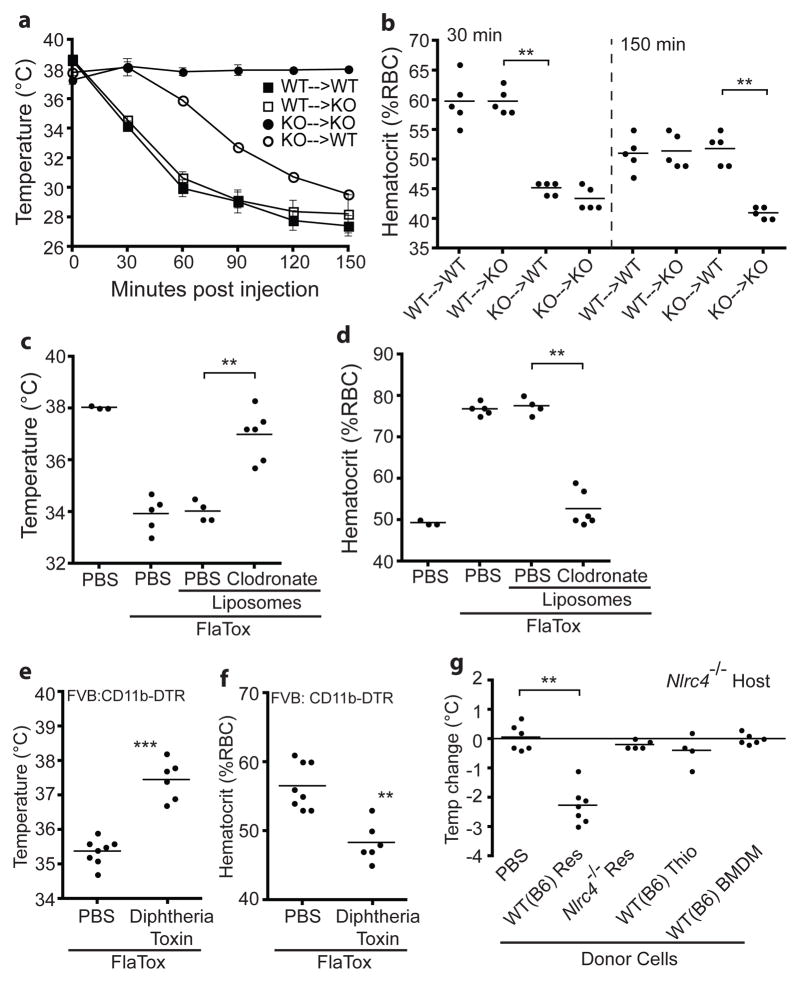
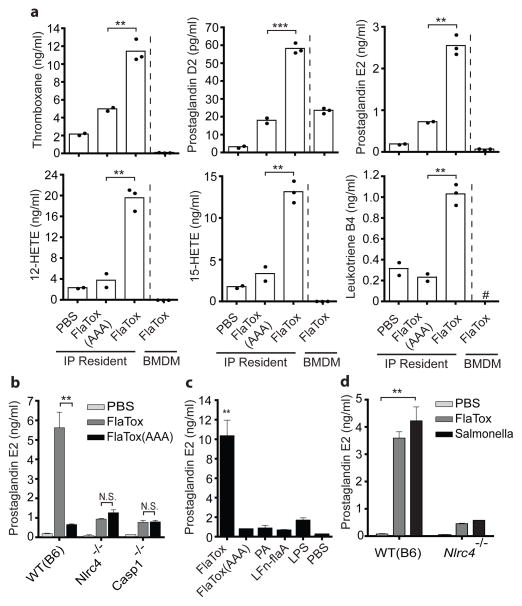
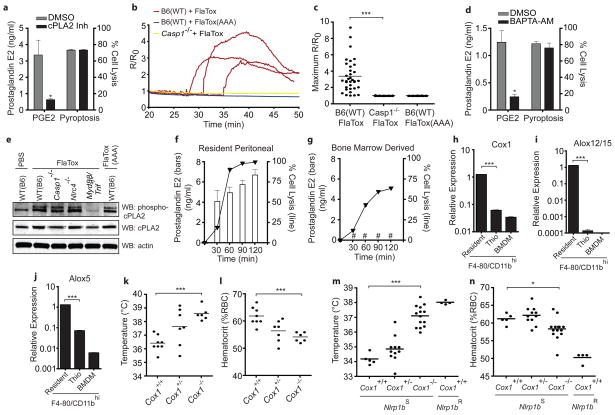
Detection of microbial products by host inflammasomes is an important mechanism of innate immune surveillance. Inflammasomes activate the caspase-1 (CASP1) protease, which processes the cytokines interleukin (IL)-1β and IL-18, and initiates a lytic host cell death called pyroptosis. To identify novel CASP1 functions in vivo, we devised a strategy for cytosolic delivery of bacterial flagellin, a specific ligand for the NAIP5 (NLR family, apoptosis inhibitory protein 5)/NLRC4 (NLR family, CARD-domain-containing 4) inflammasome. Here we show that systemic inflammasome activation by flagellin leads to a loss of vascular fluid into the intestine and peritoneal cavity, resulting in rapid (less than 30 min) death in mice. This unexpected response depends on the inflammasome components NAIP5, NLRC4 and CASP1, but is independent of the production of IL-1β or IL-18. Instead, inflammasome activation results, within minutes, in an 'eicosanoid storm'--a pathological release of signalling lipids, including prostaglandins and leukotrienes, that rapidly initiate inflammation and vascular fluid loss. Mice deficient in cyclooxygenase-1, a critical enzyme in prostaglandin biosynthesis, are resistant to these rapid pathological effects of systemic inflammasome activation by either flagellin or anthrax lethal toxin. Inflammasome-dependent biosynthesis of eicosanoids is mediated by the activation of cytosolic phospholipase A(2) in resident peritoneal macrophages, which are specifically primed for the production of eicosanoids by high expression of eicosanoid biosynthetic enzymes. Our results therefore identify eicosanoids as a previously unrecognized cell-type-specific signalling output of the inflammasome with marked physiological consequences in vivo.
Figures
References
METHOD REFERENCES
-
- Mariathasan S, et al. Differential activation of the inflammasome by caspase-1 adaptors ASC and Ipaf. Nature. 2004;430 (6996):213–218. - PubMed
-
- Li P, et al. Mice deficient in IL-1 beta-converting enzyme are defective in production of mature IL-1 beta and resistant to endotoxic shock. Cell. 1995;80 (3):401–411. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
