

NAT1/DAP5/p97 and atypical translational control in the Drosophila Circadian Oscillator
- PMID: 22904033
- PMCID: PMC3522168
- DOI: 10.1534/genetics.112.143248
NAT1/DAP5/p97 and atypical translational control in the Drosophila Circadian Oscillator
Abstract
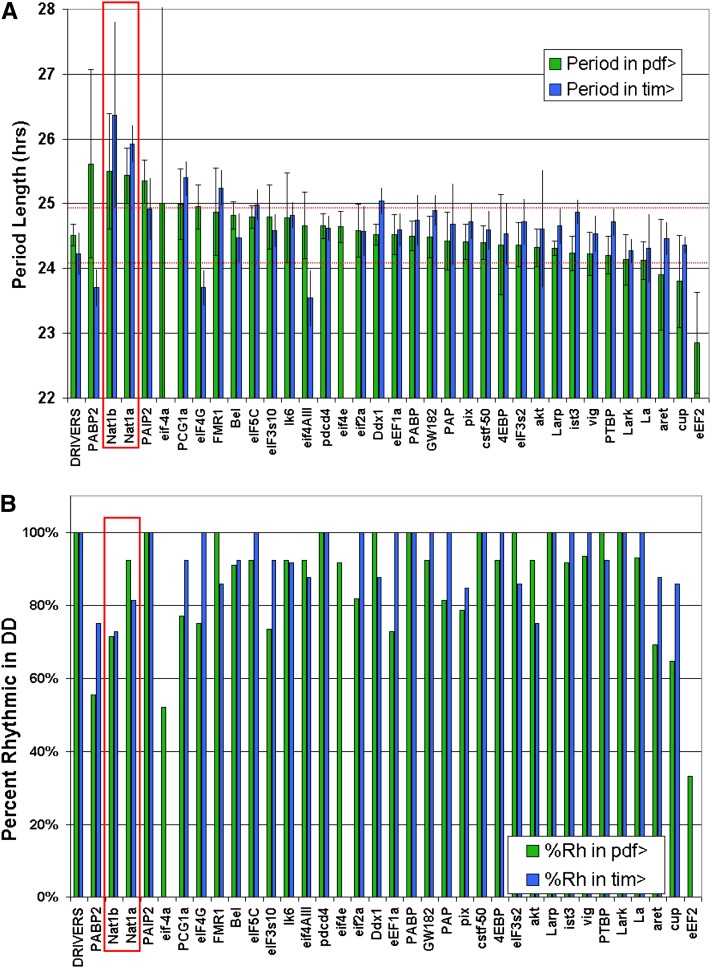
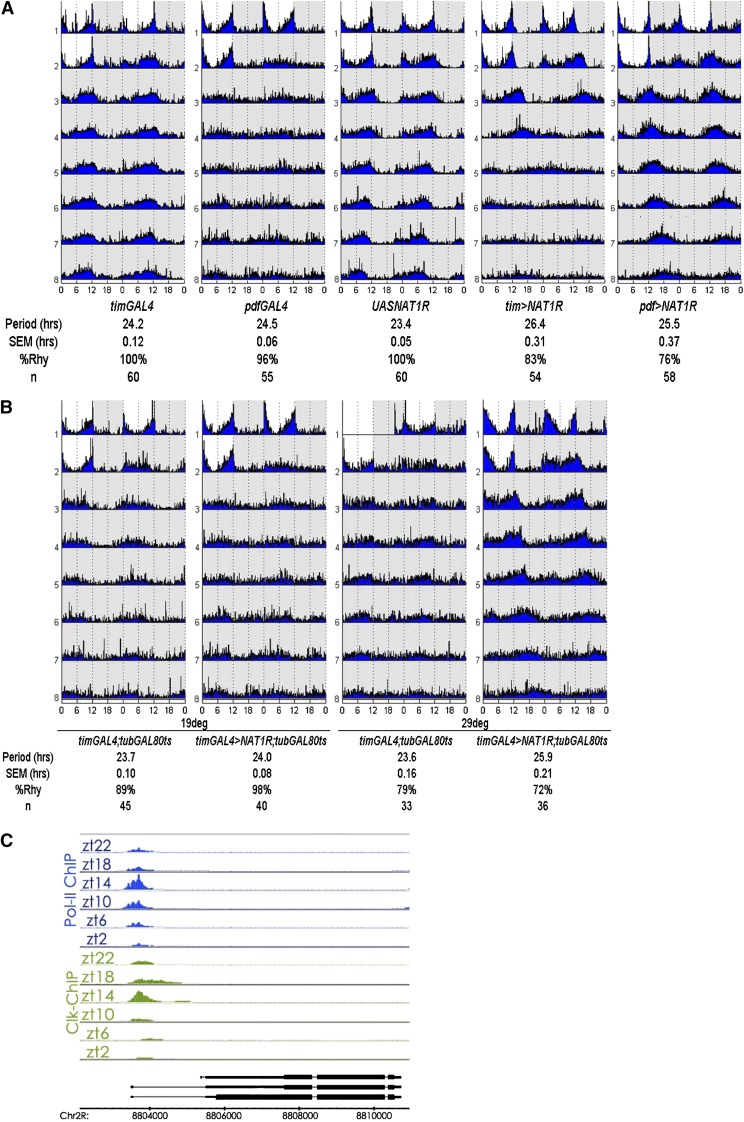
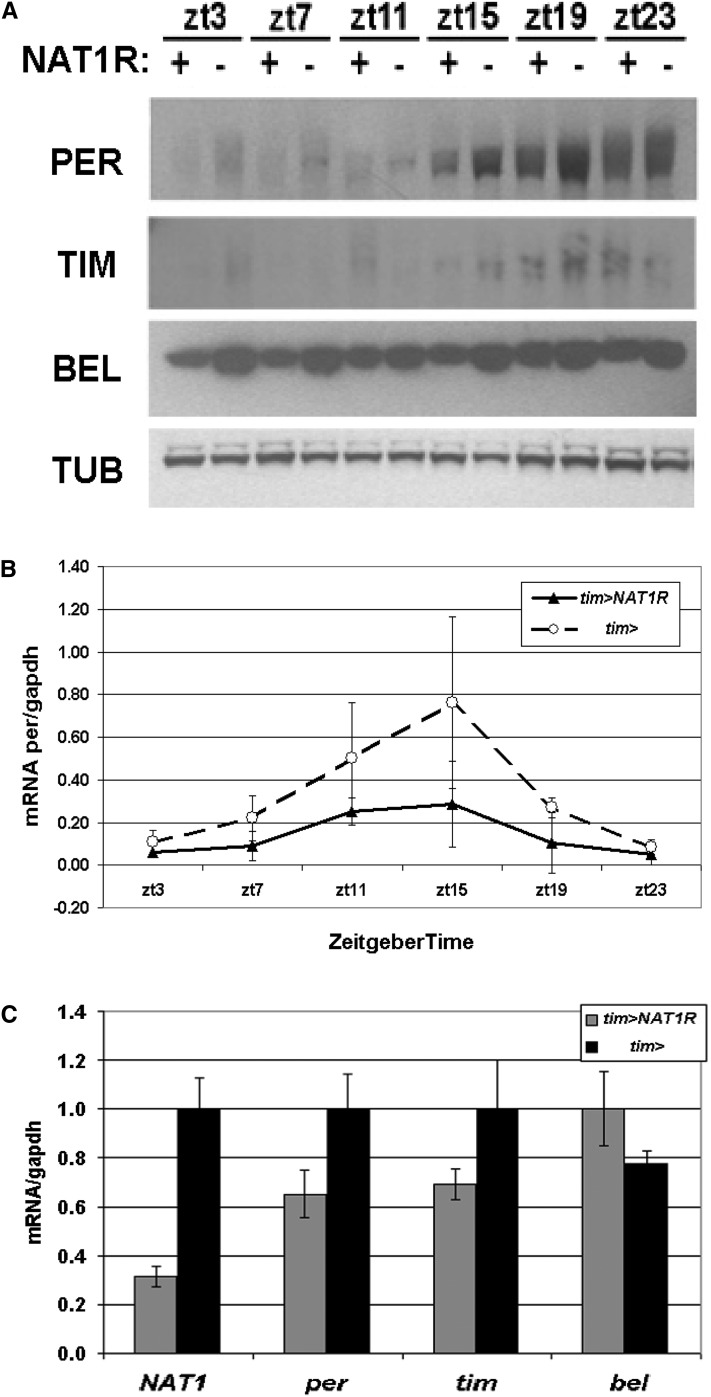
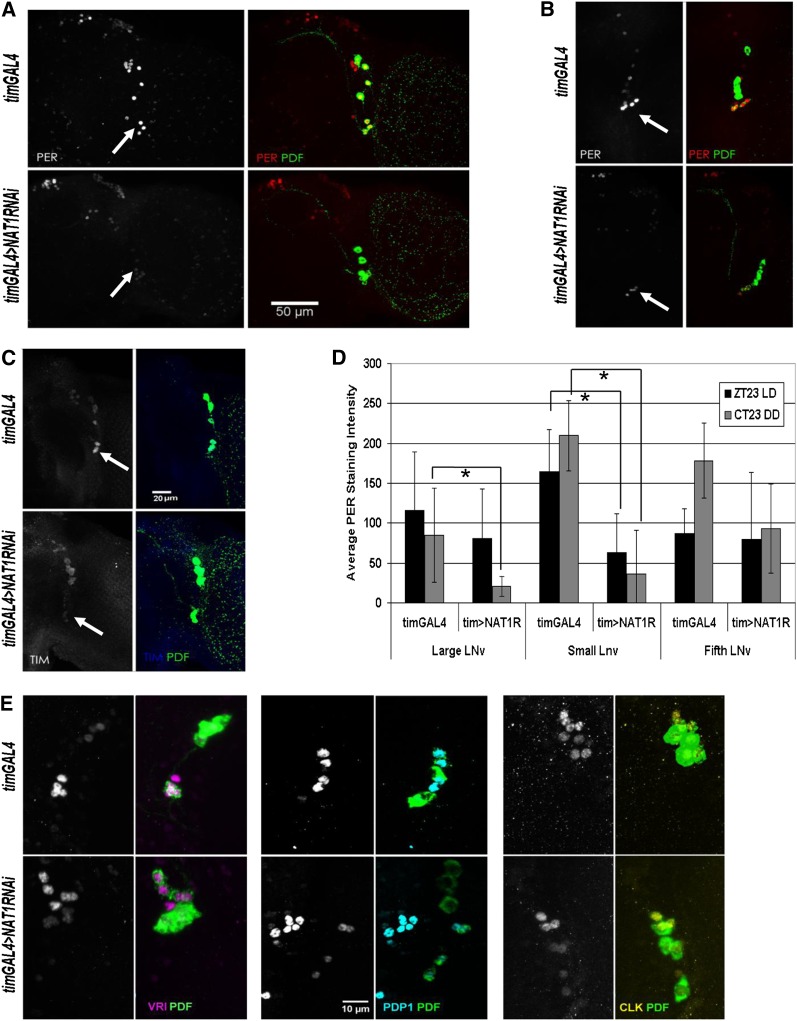
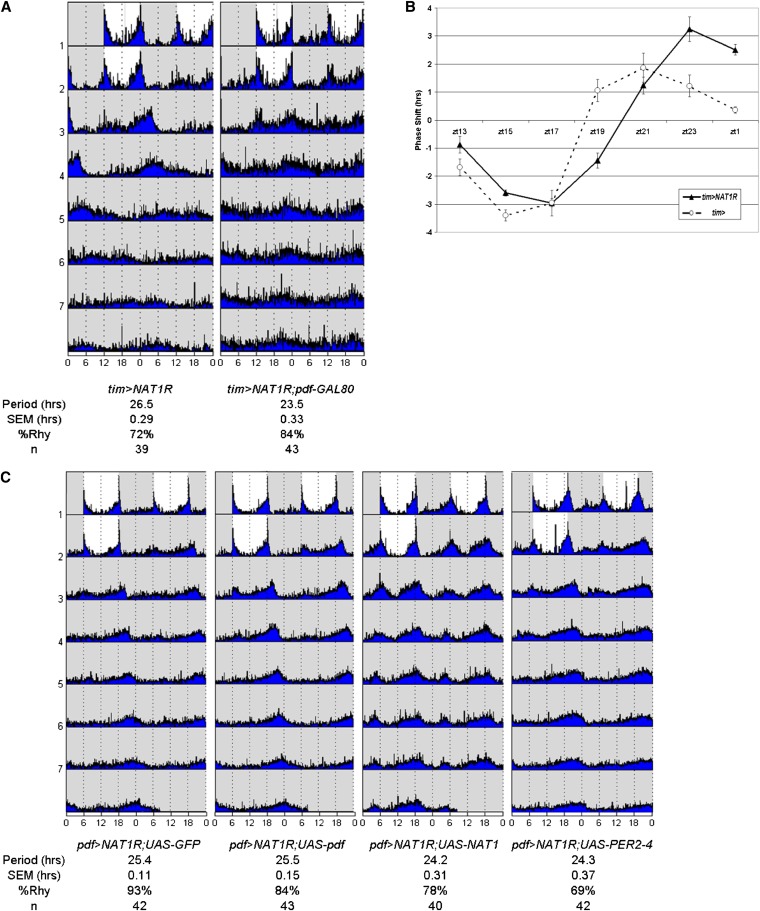
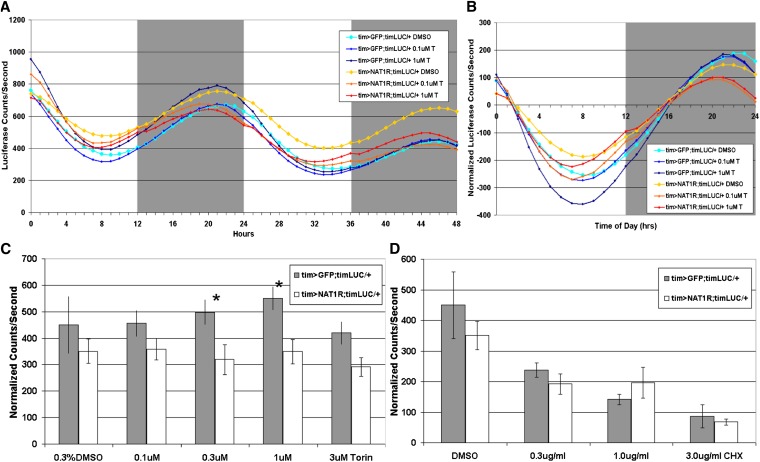
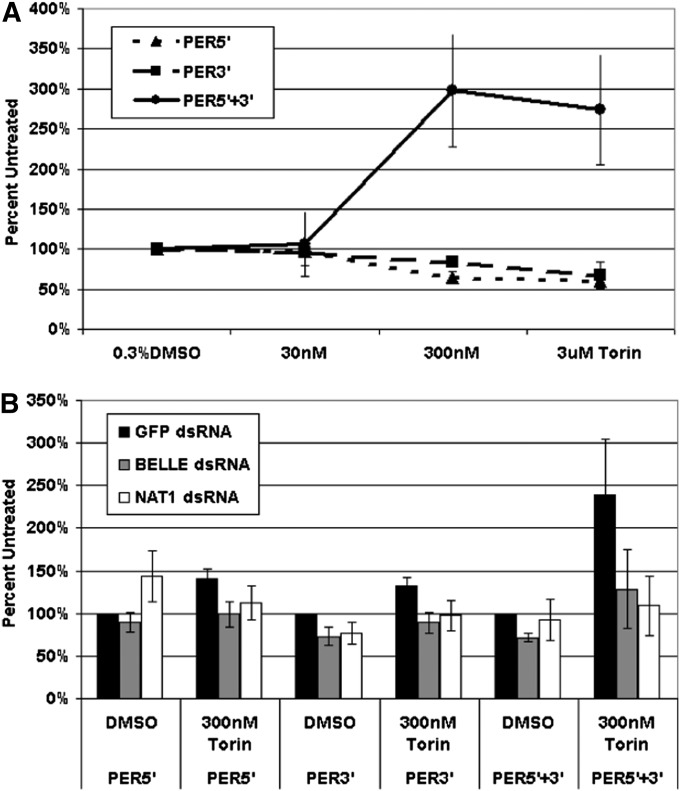
Circadian rhythms are driven by gene expression feedback loops in metazoans. Based on the success of genetic screens for circadian mutants in Drosophila melanogaster, we undertook a targeted RNAi screen to study the impact of translation control genes on circadian locomotor activity rhythms in flies. Knockdown of vital translation factors in timeless protein-positive circadian neurons caused a range of effects including lethality. Knockdown of the atypical translation factor NAT1 had the strongest effect and lengthened circadian period. It also dramatically reduced PER protein levels in pigment dispersing factor (PDF) neurons. BELLE (BEL) protein was also reduced by the NAT1 knockdown, presumably reflecting a role of NAT1 in belle mRNA translation. belle and NAT1 are also targets of the key circadian transcription factor Clock (CLK). Further evidence for a role of NAT1 is that inhibition of the target of rapamycin (TOR) kinase increased oscillator activity in cultured wings, which is absent under conditions of NAT1 knockdown. Moreover, the per 5'- and 3'-UTRs may function together to facilitate cap-independent translation under conditions of TOR inhibition. We suggest that NAT1 and cap-independent translation are important for per mRNA translation, which is also important for the circadian oscillator. A circadian translation program may be especially important in fly pacemaker cells.
Figures
References
-
- Allada R., White N. E., So W. V., Hall J. C., Rosbash M., 1998. A mutant Drosophila homolog of mammalian Clock disrupts circadian rhythms and transcription of period and timeless. Cell 93: 791–804. - PubMed
-
- Baylies M. K., Bargiello T. A., Jackson F. R., Young M. W., 1987. Changes in abundance and structure of the per gene product can alter periodicity of the Drosophila clock. Nature 326: 390–392. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
