

Predatory bacteria as natural modulators of Vibrio parahaemolyticus and Vibrio vulnificus in seawater and oysters
- PMID: 22904049
- PMCID: PMC3457099
- DOI: 10.1128/AEM.01594-12
Predatory bacteria as natural modulators of Vibrio parahaemolyticus and Vibrio vulnificus in seawater and oysters
Abstract
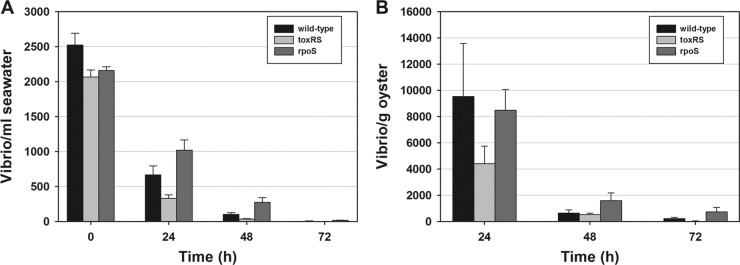
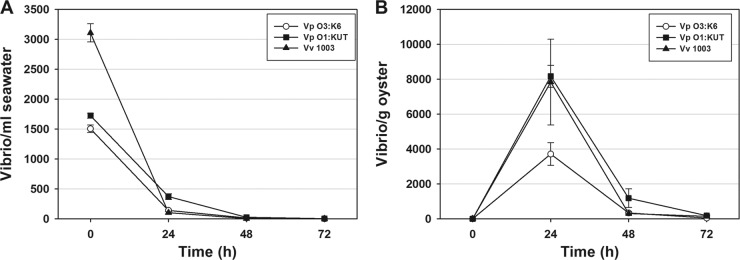
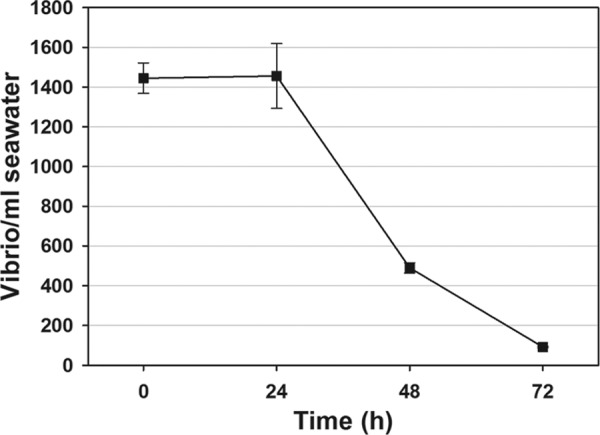
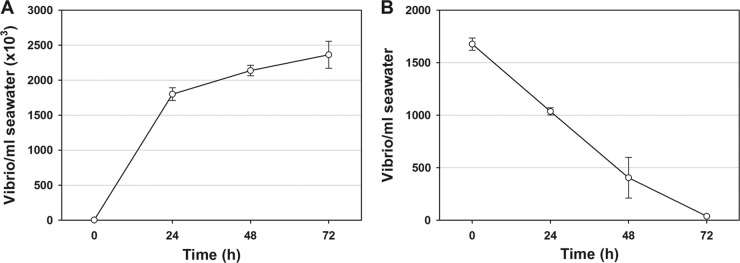
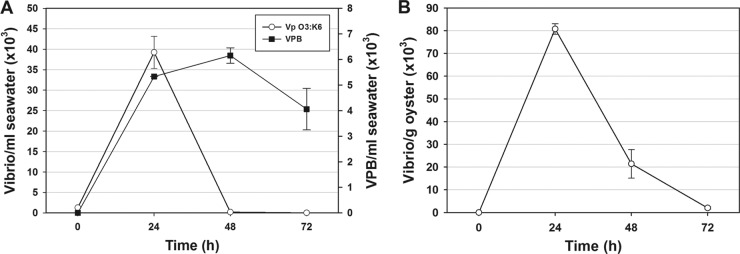
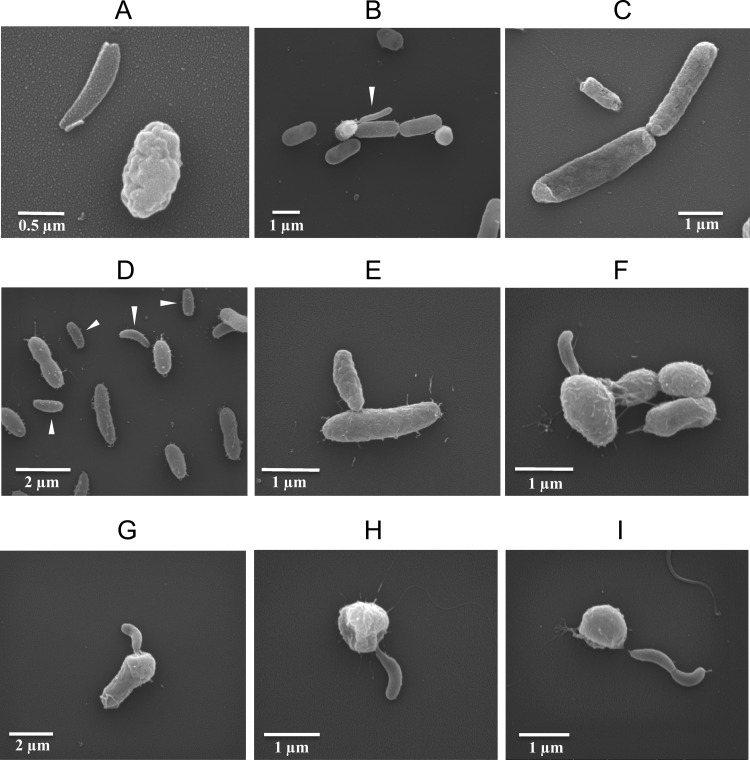
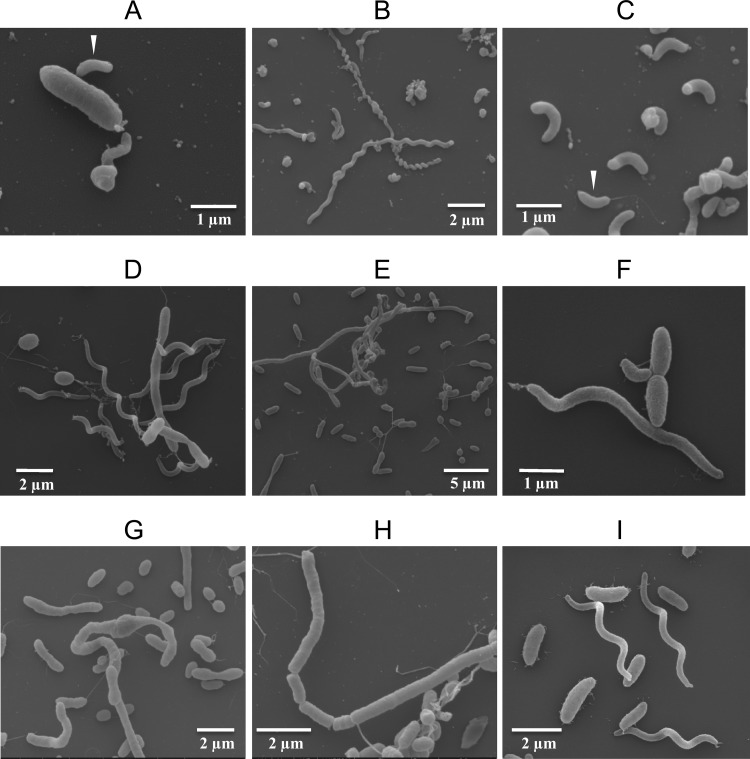
This study shows that naturally occurring Vibrio predatory bacteria (VPB) exert a major role in controlling pathogenic vibrios in seawater and shellfish. The growth and persistence of Vibrio parahaemolyticus and Vibrio vulnificus were assessed in natural seawater and in the Eastern oyster, Crassostrea virginica. The pathogens examined were V. vulnificus strain VV1003, V. parahaemolyticus O1:KUT (KUT stands for K untypeable), and V. parahaemolyticus O3:K6 and corresponding O3:K6 mutants deficient in the toxRS virulence regulatory gene or the rpoS alternative stress response sigma factor gene. Vibrios were selected for streptomycin resistance, which facilitated their enumeration. In natural seawater, oysters bioconcentrated each Vibrio strain for 24 h at 22°C; however, counts rapidly declined to near negligible levels by 72 h. In natural seawater with or without oysters, vibrios decreased more than 3 log units to near negligible levels within 72 h. Neither toxRS nor rpoS had a significant effect on Vibrio levels. In autoclaved seawater, V. parahaemolyticus O3:K6 counts increased 1,000-fold over 72 h. Failure of the vibrios to persist in natural seawater and oysters led to screening of the water samples for VPB on lawns of V. parahaemolyticus O3:K6 host cells. Many VPB, including Bdellovibrio and like organisms (BALOs; Bdellovibrio bacteriovorus and Bacteriovorax stolpii) and Micavibrio aeruginosavorus-like predators, were detected by plaque assay and electron microscopic analysis of plaque-purified isolates from Atlantic, Gulf Coast, and Hawaiian seawater. When V. parahaemolyticus O3:K6 was added to natural seawater containing trace amounts of VPB, Vibrio counts diminished 3 log units to nondetectable levels, while VPB increased 3 log units within 48 h. We propose a new paradigm that VPB are important modulators of pathogenic vibrios in seawater and oysters.
Figures
References
-
- Centers for Disease Control and Prevention 1999. Outbreak of Vibrio parahaemolyticus infection associated with eating raw oysters and clams harvested from Long Island Sound - Connecticut, New Jersey, and New York, 1998. MMWR Morb. Mortal. Wkly. Rep. 48:48–51 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
