

Inhibition of JAKs in macrophages increases lipopolysaccharide-induced cytokine production by blocking IL-10-mediated feedback
- PMID: 22904308
- PMCID: PMC3443740
- DOI: 10.4049/jimmunol.1200310
Inhibition of JAKs in macrophages increases lipopolysaccharide-induced cytokine production by blocking IL-10-mediated feedback
Abstract
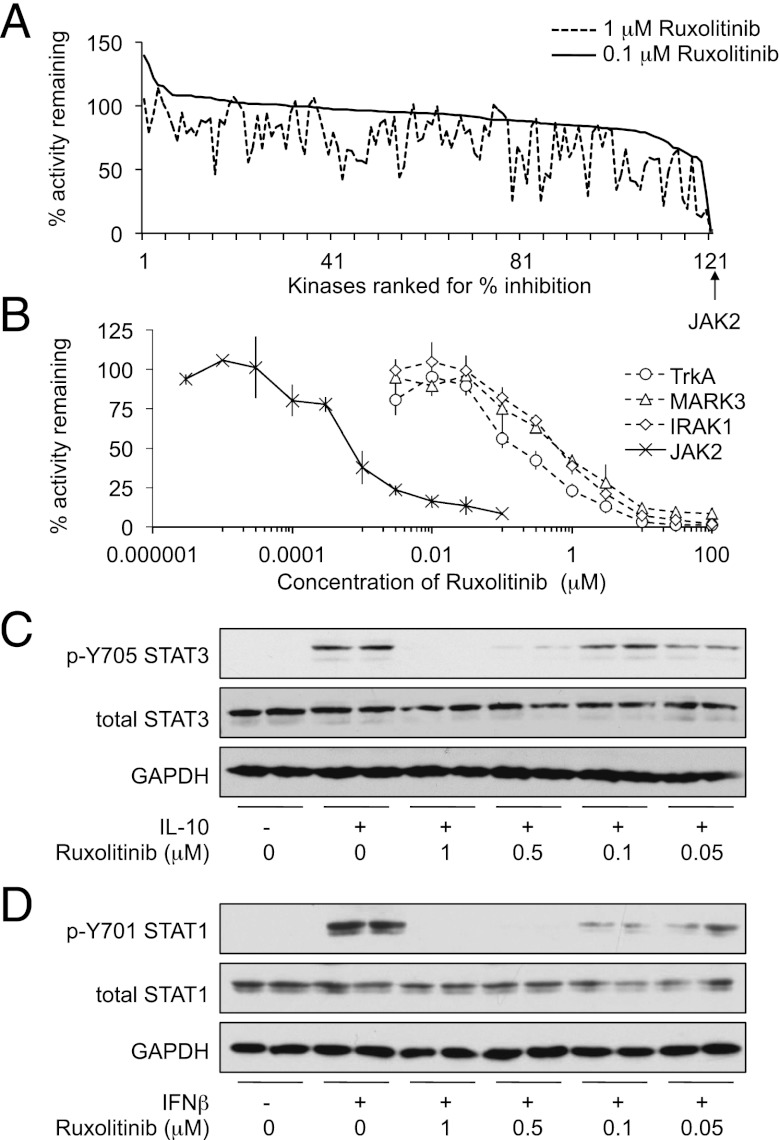
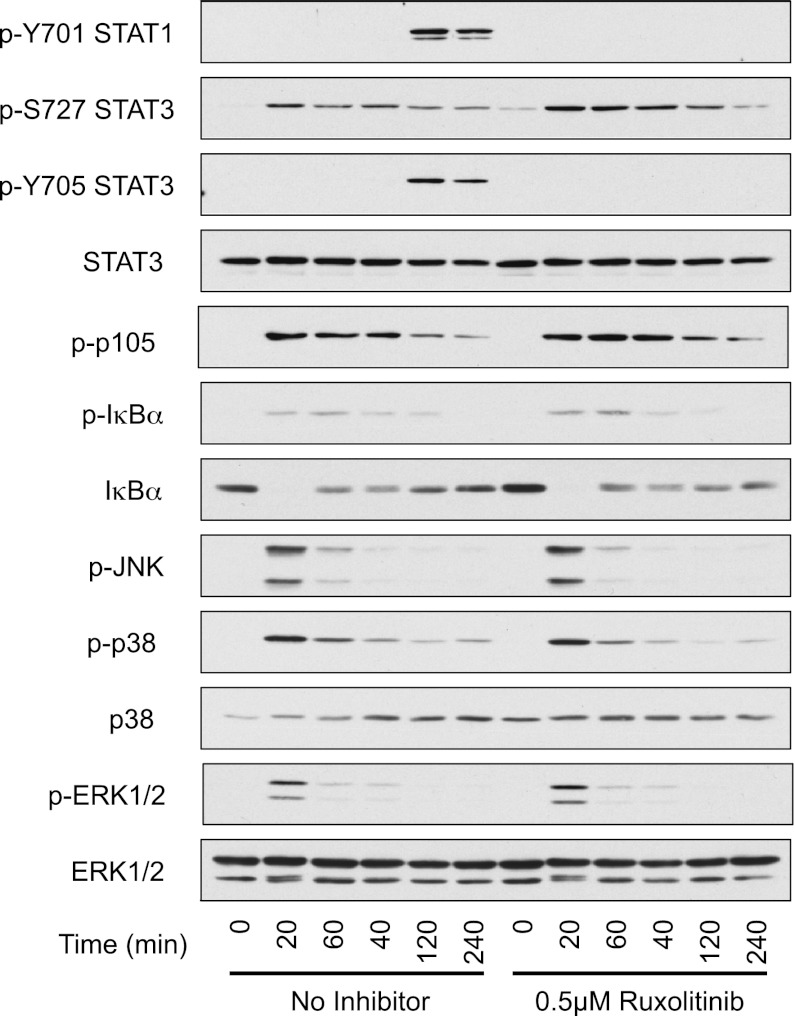
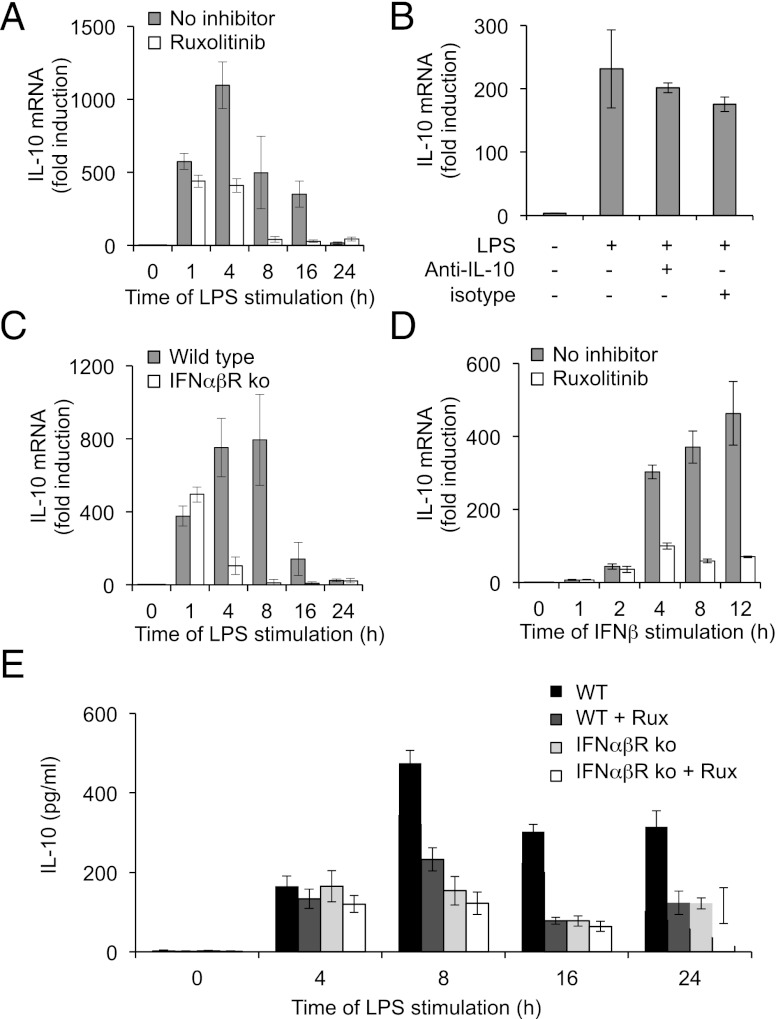
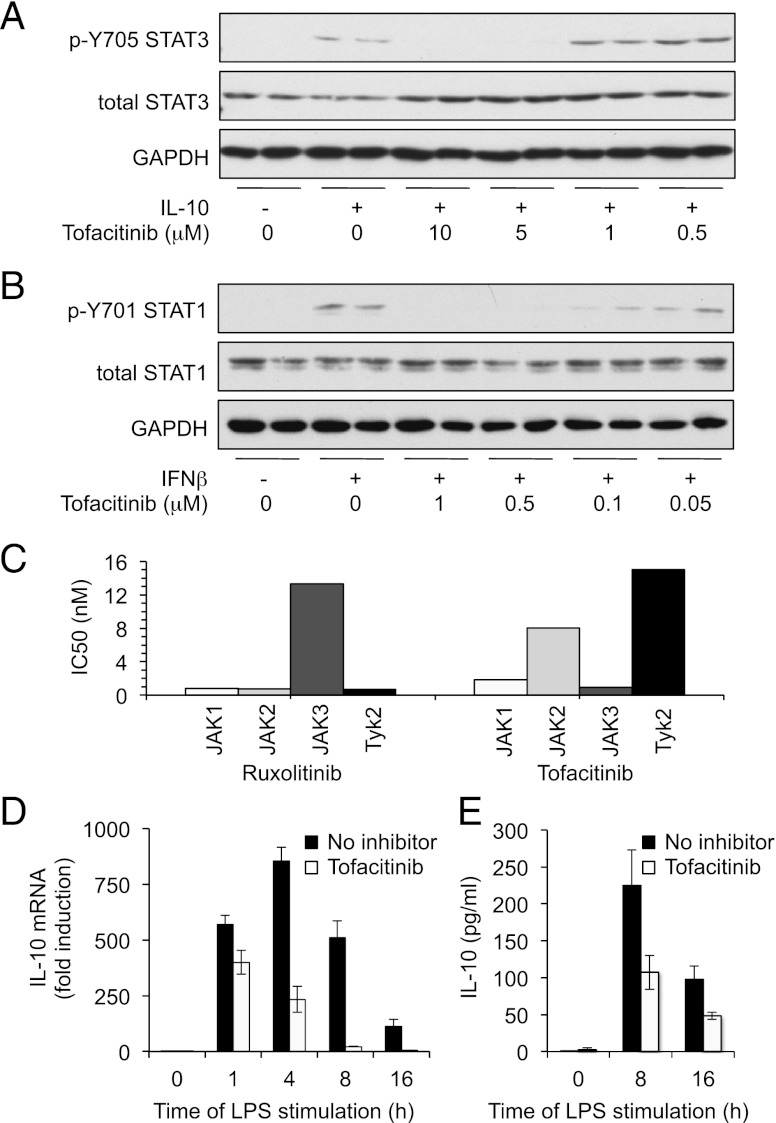
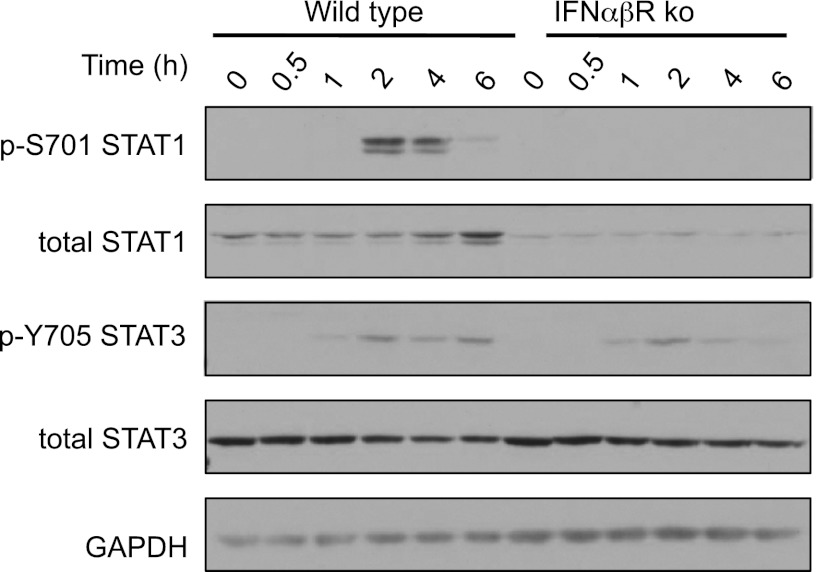
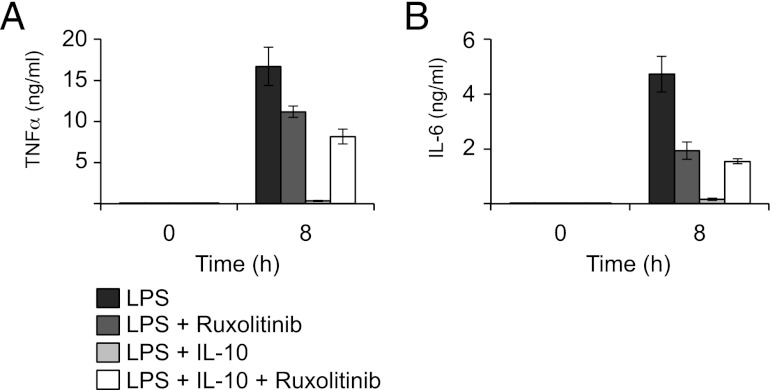
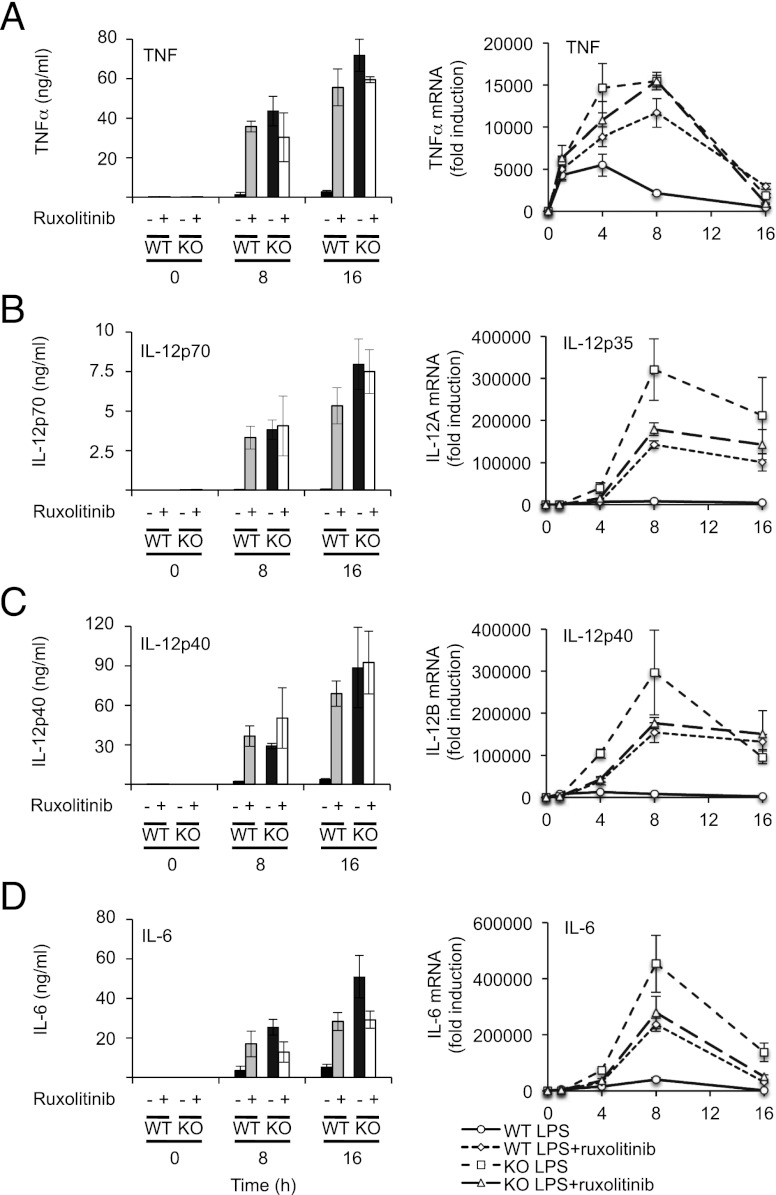
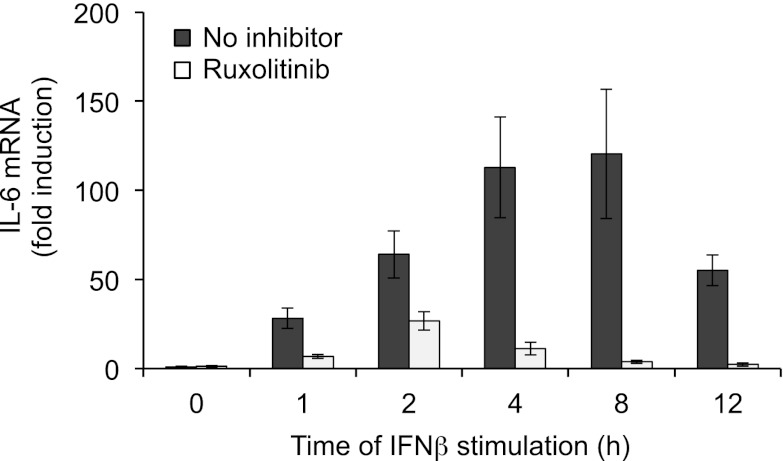
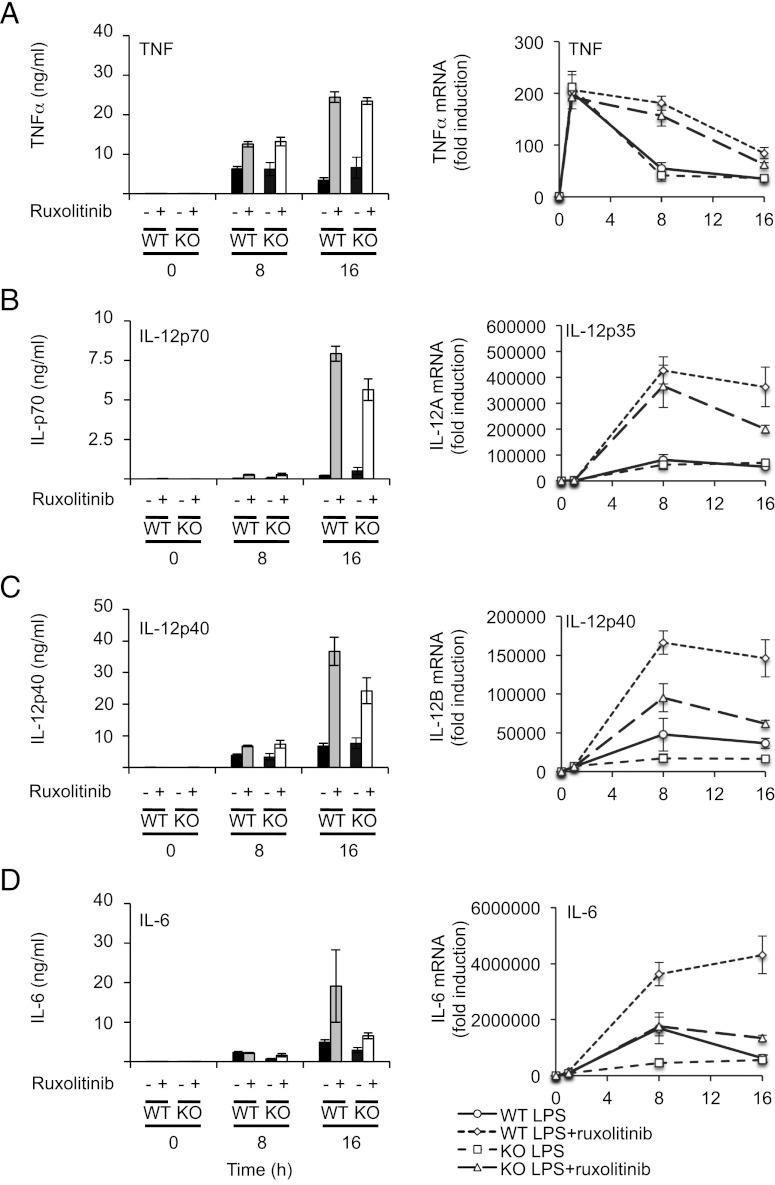
Macrophages are an important source of cytokines following infection. Stimulation of macrophages with TLR agonists results in the secretion of TNF-α, IL-6, and IL-12, and the production of these cytokines is controlled by multiple feedback pathways. Macrophages also produce IL-10, which acts to inhibit proinflammatory cytokine production by macrophages via a JAK/STAT3-dependent pathway. We show in this paper that, Ruxolitinib, a recently described selective inhibitor of JAKs, increases TNF, IL-6, and IL-12 secretion in mouse bone marrow-derived macrophages stimulated with LPS. This effect is largely due to its ability to block IL-10-mediated feedback inhibition on cytokine transcription in macrophages. Similar results were also obtained with a second structurally unrelated Jak inhibitor, Tofacitinib. In addition, LPS induced the production of IFN-β, which was then able to activate JAKs in macrophages, resulting in the stimulation of STAT1 phosphorylation. The initial induction of IL-10 was independent of JAK signaling; however, inhibition of JAKs did reduce IL-10 secretion at later time points. This reflected a requirement for the IFN-β feedback loop to sustain IL-10 transcription following LPS stimulation. In addition to IL-10, IFN-β also helped sustain IL-6 and IL-12 transcription. Overall, these results suggest that inhibition of JAKs may increase the inflammatory potential of macrophages stimulated with TLR4 agonists.
Figures
References
-
- Schindler C., Levy D. E., Decker T. 2007. JAK-STAT signaling: from interferons to cytokines. J. Biol. Chem. 282: 20059–20063 - PubMed
-
- Levy D. E., Darnell J. E., Jr. 2002. Stats: transcriptional control and biological impact. Nat. Rev. Mol. Cell Biol. 3: 651–662 - PubMed
-
- Quintás-Cardama A., Kantarjian H., Cortes J., Verstovsek S. 2011. Janus kinase inhibitors for the treatment of myeloproliferative neoplasias and beyond. Nat. Rev. Drug Discov. 10: 127–140 - PubMed
-
- Vijayakrishnan L., Venkataramanan R., Gulati P. 2011. Treating inflammation with the Janus kinase inhibitor CP-690,550. Trends Pharmacol. Sci. 32: 25–34 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
