

Variation in infectivity and aggressiveness in space and time in wild host-pathogen systems: causes and consequences
- PMID: 22905782
- PMCID: PMC3440529
- DOI: 10.1111/j.1420-9101.2012.02588.x
Variation in infectivity and aggressiveness in space and time in wild host-pathogen systems: causes and consequences
Abstract
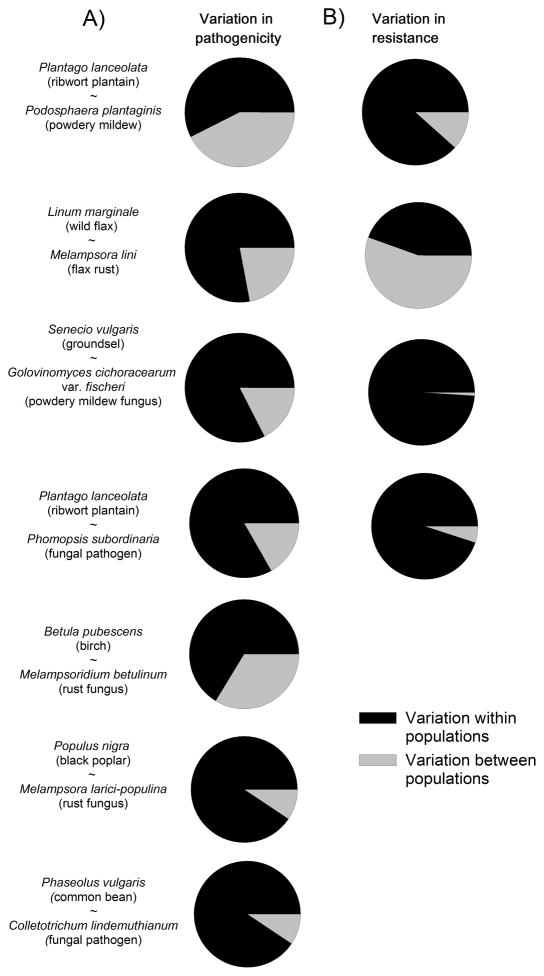
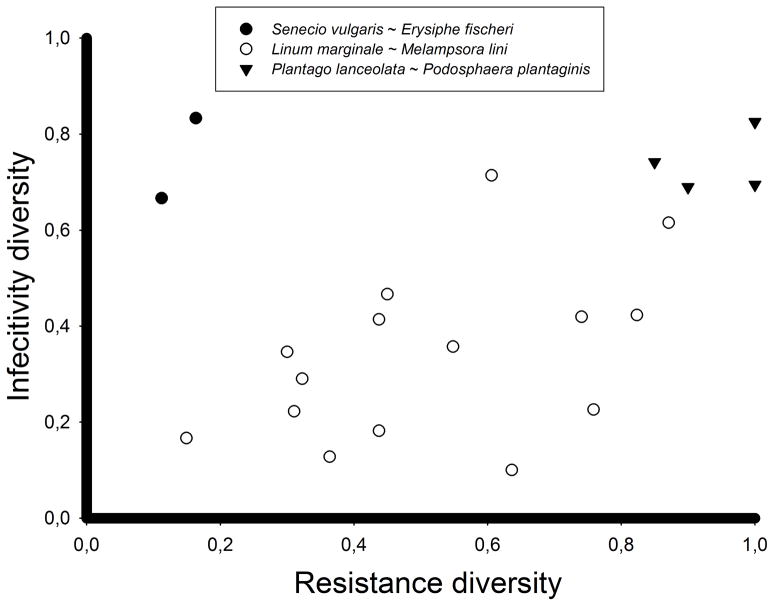
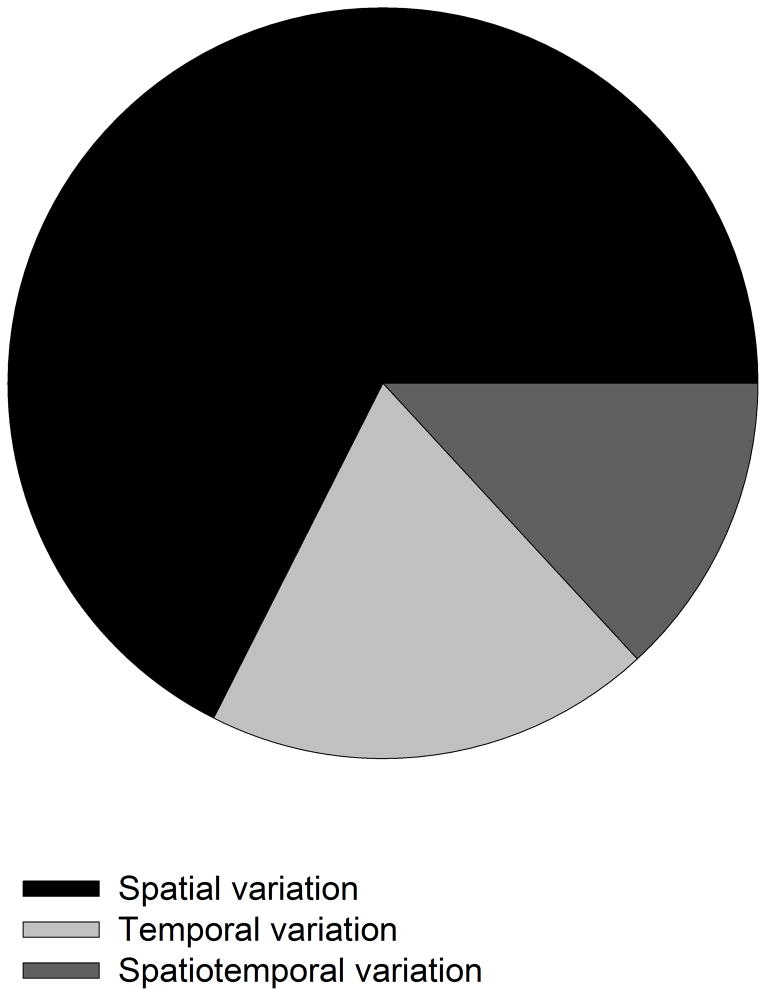
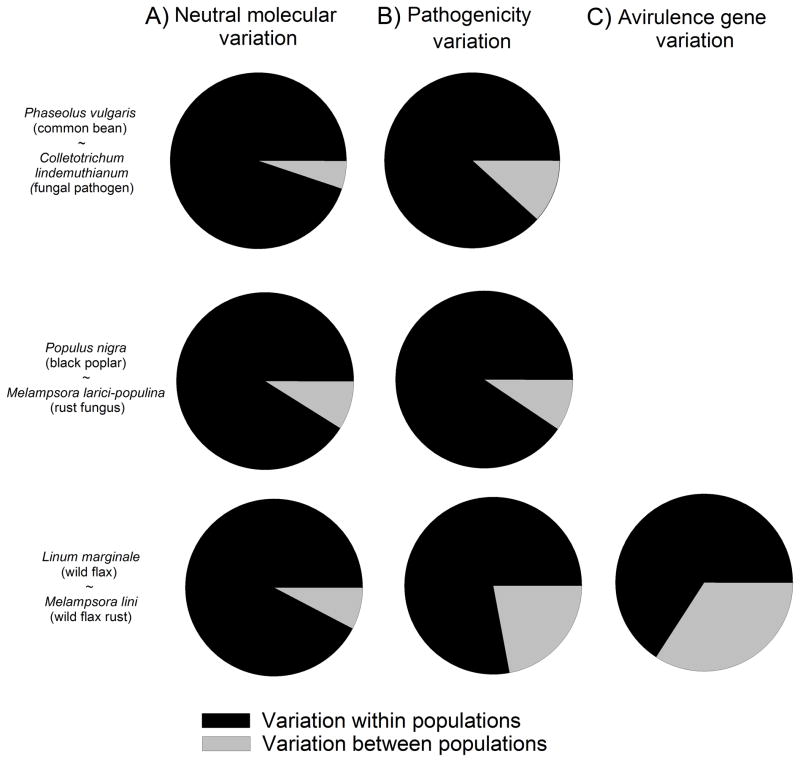
Variation in host resistance and in the ability of pathogens to infect and grow (i.e. pathogenicity) is important as it provides the raw material for antagonistic (co)evolution and therefore underlies risks of disease spread, disease evolution and host shifts. Moreover, the distribution of this variation in space and time may inform us about the mode of coevolutionary selection (arms race vs. fluctuating selection dynamics) and the relative roles of G × G interactions, gene flow, selection and genetic drift in shaping coevolutionary processes. Although variation in host resistance has recently been reviewed, little is known about overall patterns in the frequency and scale of variation in pathogenicity, particularly in natural systems. Using 48 studies from 30 distinct host-pathogen systems, this review demonstrates that variation in pathogenicity is ubiquitous across multiple spatial and temporal scales. Quantitative analysis of a subset of extensively studied plant-pathogen systems shows that the magnitude of within-population variation in pathogenicity is large relative to among-population variation and that the distribution of pathogenicity partly mirrors the distribution of host resistance. At least part of the variation in pathogenicity found at a given spatial scale is adaptive, as evidenced by studies that have examined local adaptation at scales ranging from single hosts through metapopulations to entire continents and - to a lesser extent - by comparisons of pathogenicity with neutral genetic variation. Together, these results support coevolutionary selection through fluctuating selection dynamics. We end by outlining several promising directions for future research.
© 2012 The Authors. Journal of Evolutionary Biology © 2012 European Society For Evolutionary Biology.
Figures
References
-
- Abang MM, Baum M, Ceccarelli S, Grando S, Linde CC, Yahyaoui A, et al. Differential selection on Rhynchosporium secalis during parasitic and saprophytic phases in the barley scald disease cycle. Phytopathology. 2006;96:1214–1222. - PubMed
-
- Agrawal A, Lively CM. Infection genetics: gene-for-gene versus matching-alleles models and all points in between. Evol Ecol Res. 2002;4:79–90.
-
- Agrios GN. Plant pathology. 5. Academic press; New York, USA: 2005.
-
- Al-Yaman F, Genton B, Reeder JC, Anders RF, Smith T, Alpers MP. Reduced risk of clinical malaria in children infected with multiple clones of Plasmodium falciparum in a highly endemic area: a prospective community study. Trans R Soc Trop Med Hyg. 1997;91:602–605. - PubMed
-
- Alexander HM. An experimental field study of anther-smut disease of Silene alba caused by Ustilago violacea: genotypic variation and disease incidence. Evolution. 1989;43:835–847. - PubMed
