

piRNA biogenesis during adult spermatogenesis in mice is independent of the ping-pong mechanism
- PMID: 22907665
- PMCID: PMC3463270
- DOI: 10.1038/cr.2012.120
piRNA biogenesis during adult spermatogenesis in mice is independent of the ping-pong mechanism
Abstract
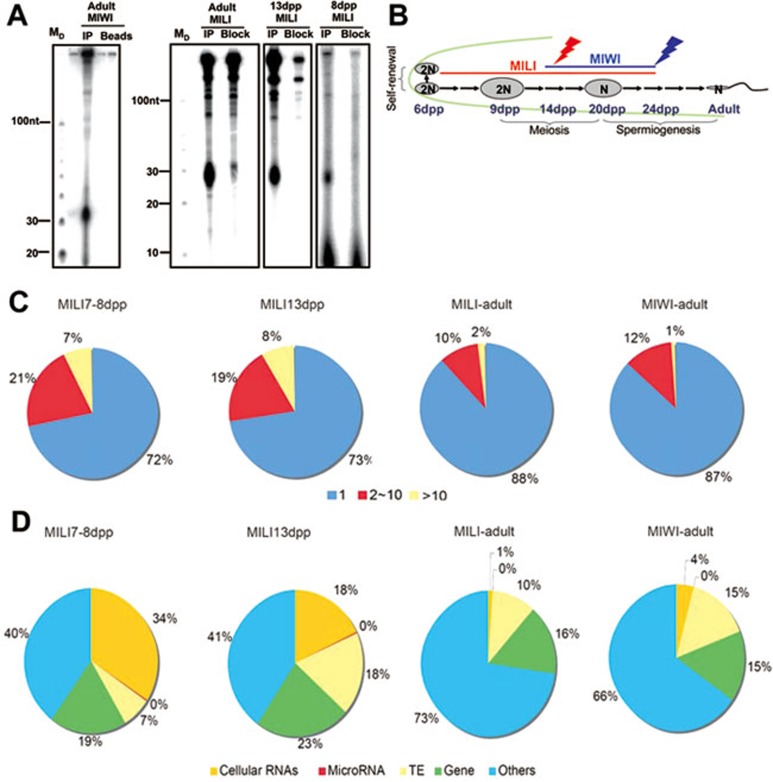
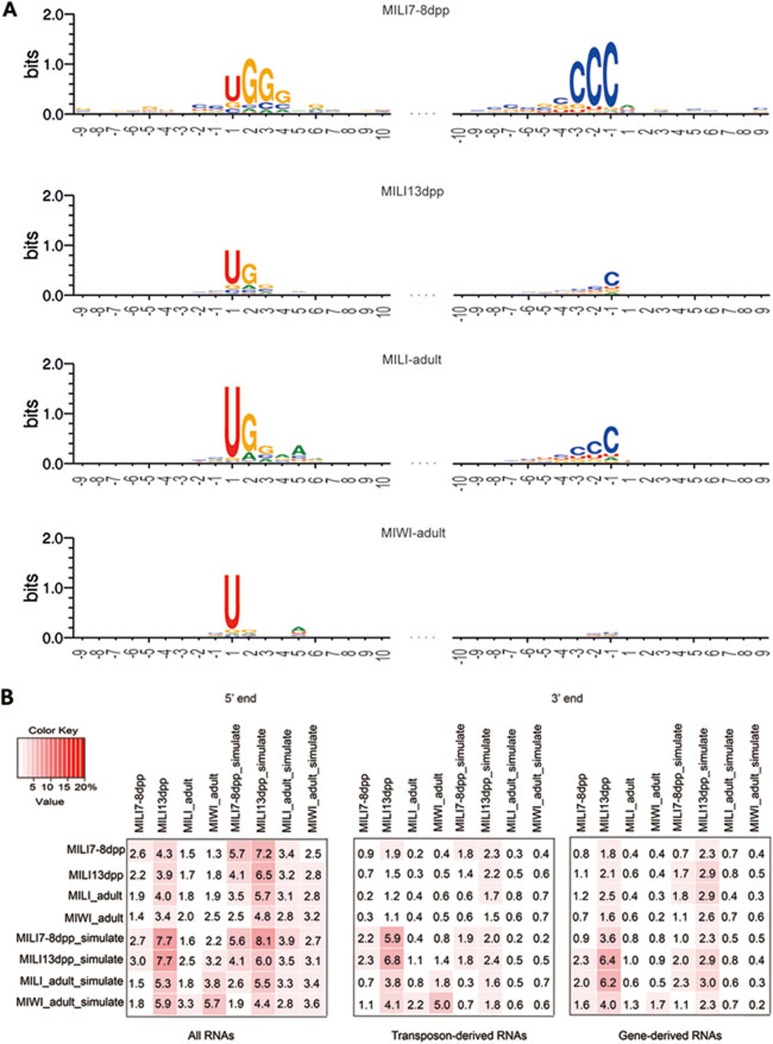
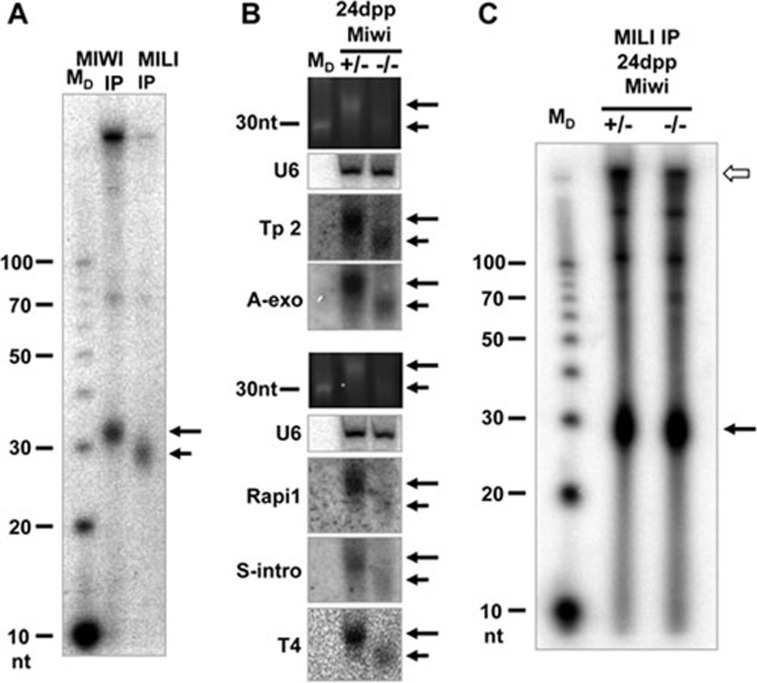
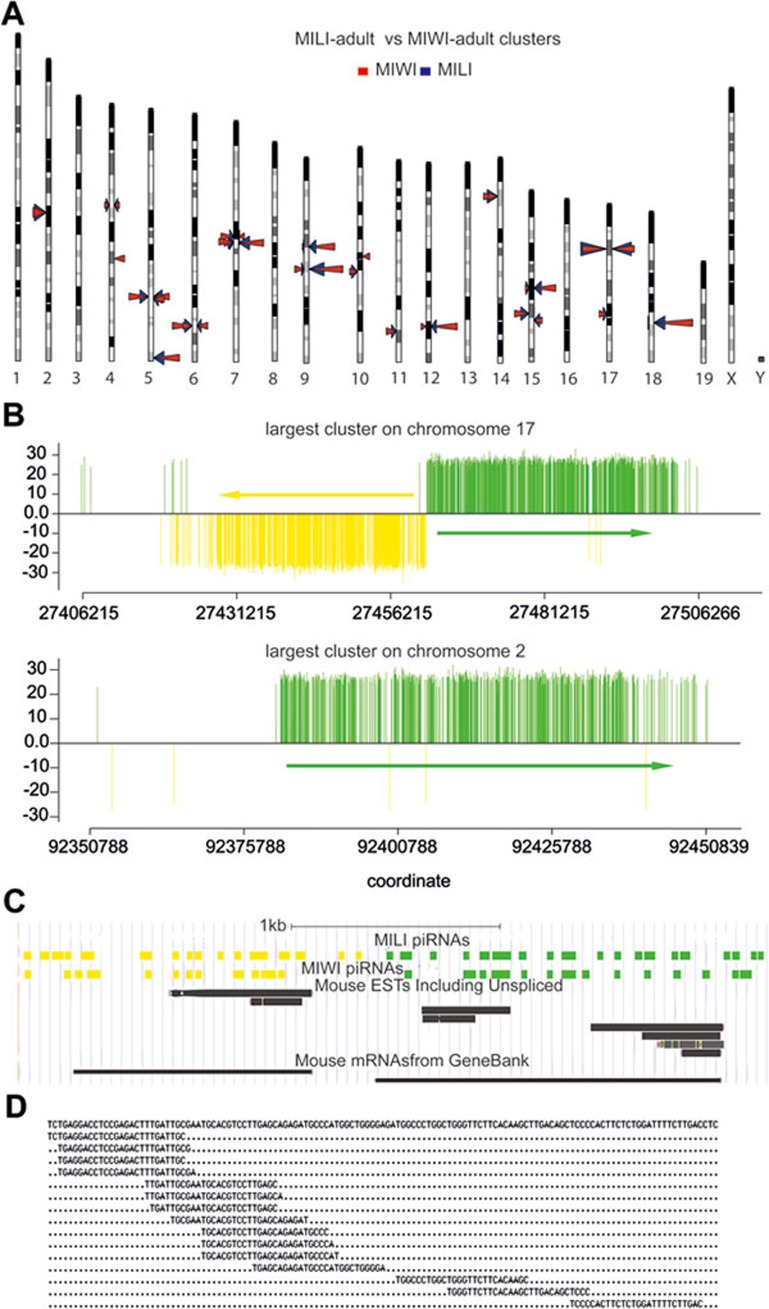
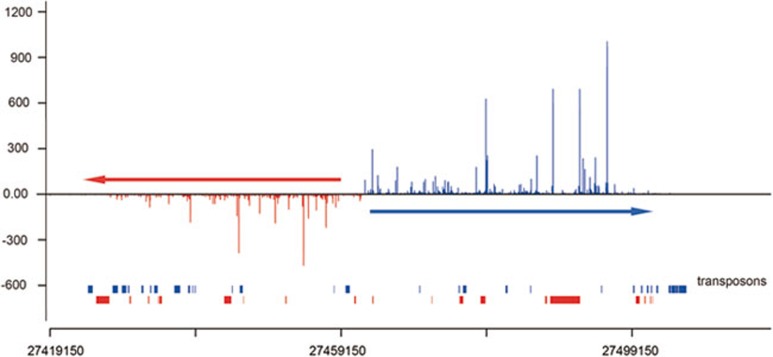
piRNAs, a class of small non-coding RNAs associated with PIWI proteins, have broad functions in germline development, transposon silencing, and epigenetic regulation. In diverse organisms, a subset of piRNAs derived from repeat sequences are produced via the interplay between two PIWI proteins. This mechanism, termed "ping-pong" cycle, operates among the PIWI proteins of the primordial mouse testis; however, its involvement in postnatal testes remains elusive. Here we show that adult testicular piRNAs are produced independent of the ping-pong mechanism. We identified and characterized large populations of piRNAs in the adult and postnatal developing testes associated with MILI and MIWI, the only PIWI proteins detectable in these testes. No interaction between MILI and MIWI or sequence feature for the ping-pong mechanism among their piRNAs was detected in the adult testis. The majority of MILI- and MIWI-associated piRNAs originate from the same DNA strands within the same loci. Both populations of piRNAs are biased for 5' Uracil but not for Adenine on the 10th nucleotide position, and display no complementarity. Furthermore, in Miwi mutants, MILI-associated piRNAs are not downregulated, but instead upregulated. These results indicate that the adult testicular piRNAs are predominantly, if not exclusively, produced by a primary processing mechanism instead of the ping-pong mechanism. In this primary pathway, biogenesis of MILI- and MIWI-associated piRNAs may compete for the same precursors; the types of piRNAs produced tend to be non-selectively dictated by the available precursors in the cell; and precursors with introns tend to be spliced before processed into piRNAs.
Figures
References
-
- Brennecke J, Aravin AA, Stark A, et al. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell. 2007;128:1089–1103. - PubMed
-
- Gunawardane LS, Saito K, Nishida KM, et al. A slicer-mediated mechanism for repeat-associated siRNA 5′ end formation in Drosophila. Science. 2007;315:1587–1590. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
