

Myosin IC generates power over a range of loads via a new tension-sensing mechanism
- PMID: 22908250
- PMCID: PMC3443183
- DOI: 10.1073/pnas.1207811109
Myosin IC generates power over a range of loads via a new tension-sensing mechanism
Abstract
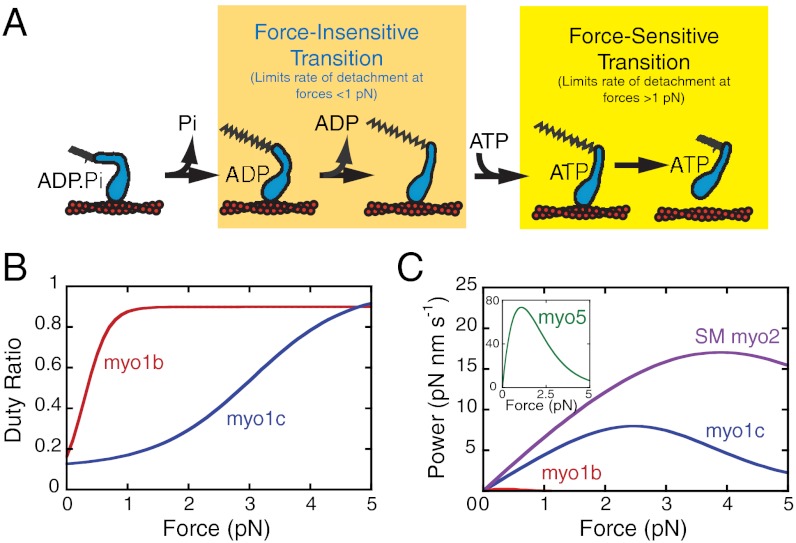
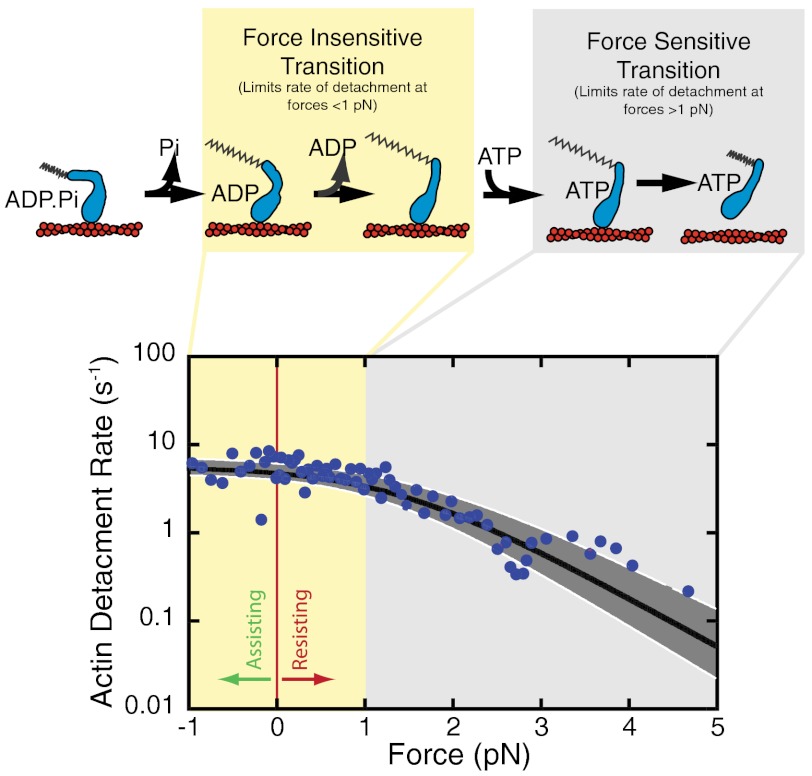
Myosin IC (myo1c), a widely expressed motor protein that links the actin cytoskeleton to cell membranes, has been associated with numerous cellular processes, including insulin-stimulated transport of GLUT4, mechanosensation in sensory hair cells, endocytosis, transcription of DNA in the nucleus, exocytosis, and membrane trafficking. The molecular role of myo1c in these processes has not been defined, so to better understand myo1c function, we utilized ensemble kinetic and single-molecule techniques to probe myo1c's biochemical and mechanical properties. Utilizing a myo1c construct containing the motor and regulatory domains, we found the force dependence of the actin-attachment lifetime to have two distinct regimes: a force-independent regime at forces < 1 pN, and a highly force-dependent regime at higher loads. In this force-dependent regime, forces that resist the working stroke increase the actin-attachment lifetime. Unexpectedly, the primary force-sensitive transition is the isomerization that follows ATP binding, not ADP release as in other slow myosins. This force-sensing behavior is unique amongst characterized myosins and clearly demonstrates mechanochemical diversity within the myosin family. Based on these results, we propose that myo1c functions as a slow transporter rather than a tension-sensitive anchor.
Conflict of interest statement
The authors declare no conflict of interest.
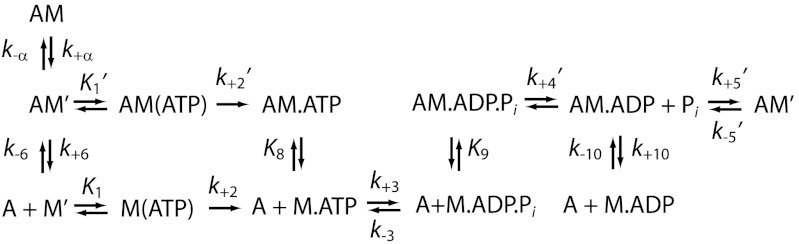
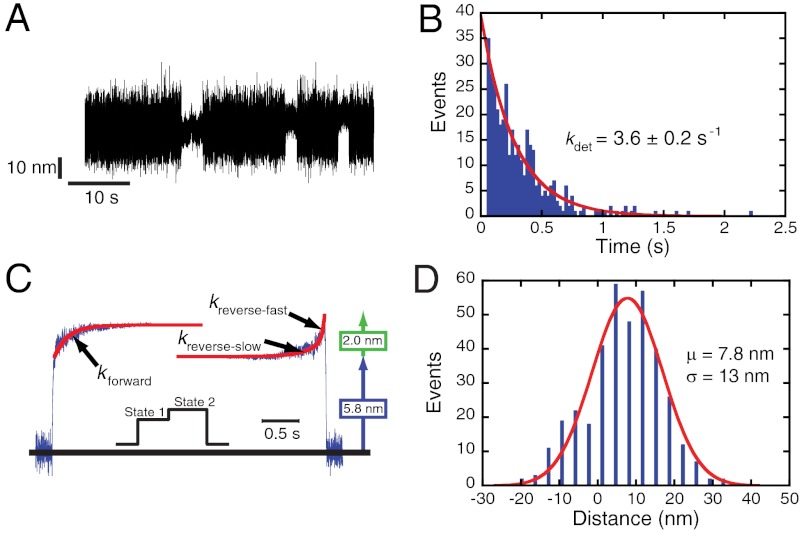
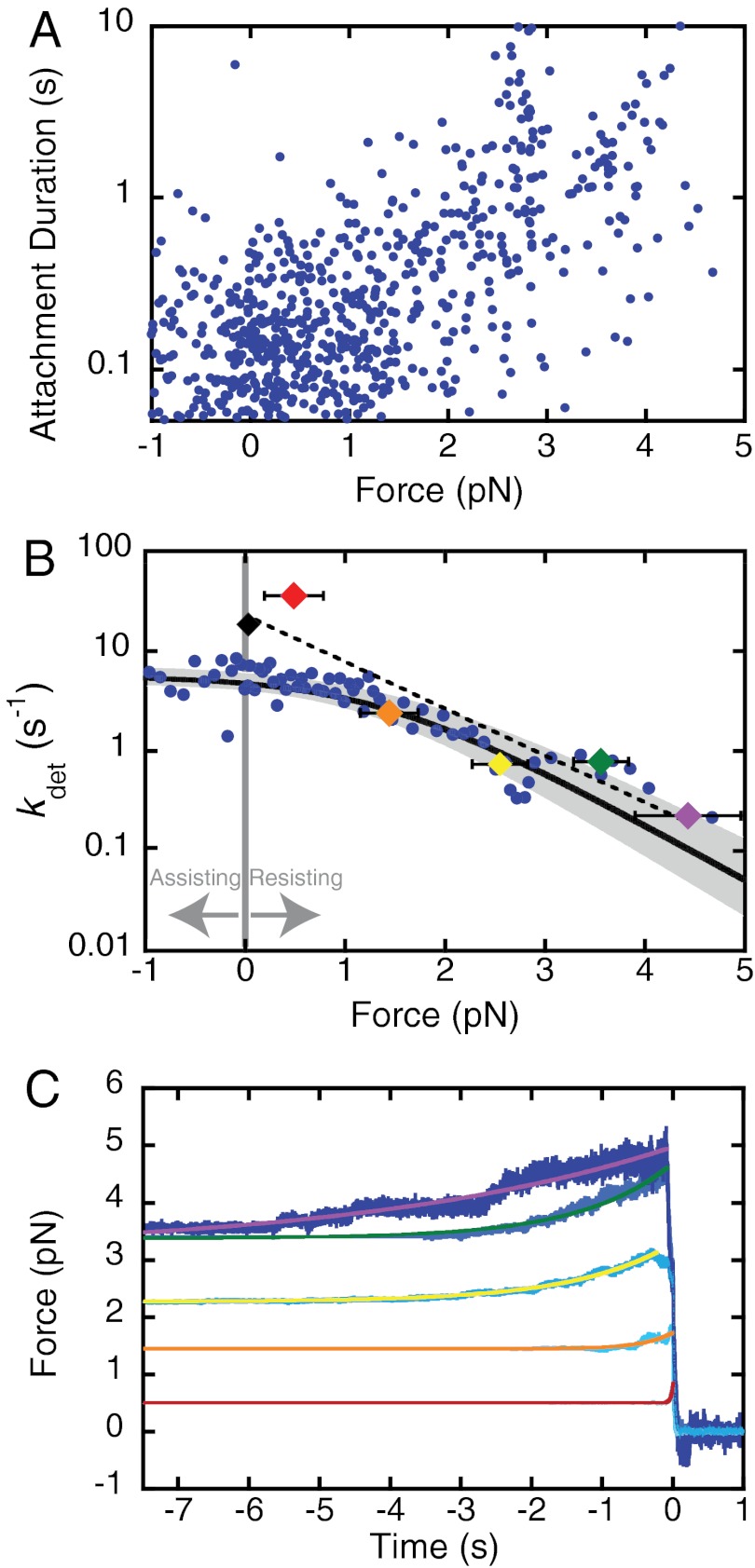
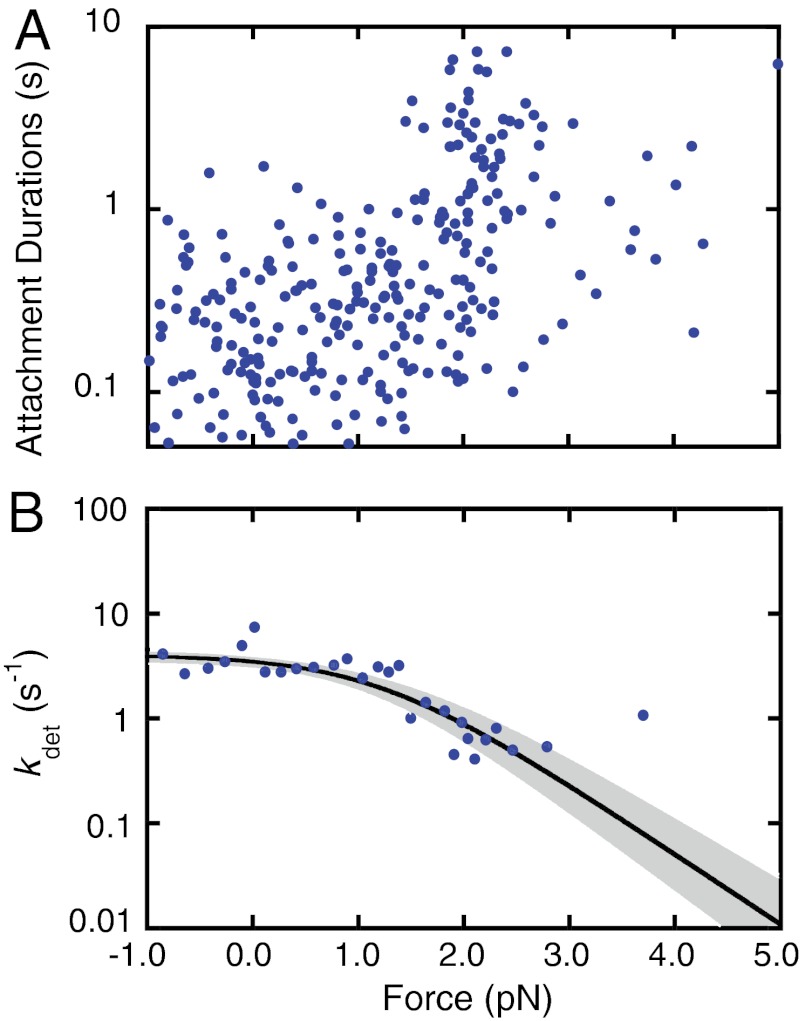
Figures
References
-
- Bose A, et al. Glucose transporter recycling in response to insulin is facilitated by myosin Myo1c. Nature. 2002;420:821–824. - PubMed
-
- Chen XW, Leto D, Chiang SH, Wang Q, Saltiel AR. Activation of RalA is required for insulin-stimulated Glut4 trafficking to the plasma membrane via the exocyst and the motor protein Myo1c. Dev Cell. 2007;13:391–404. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
