

FERONIA receptor kinase pathway suppresses abscisic acid signaling in Arabidopsis by activating ABI2 phosphatase
- PMID: 22908257
- PMCID: PMC3437822
- DOI: 10.1073/pnas.1212547109
FERONIA receptor kinase pathway suppresses abscisic acid signaling in Arabidopsis by activating ABI2 phosphatase
Abstract
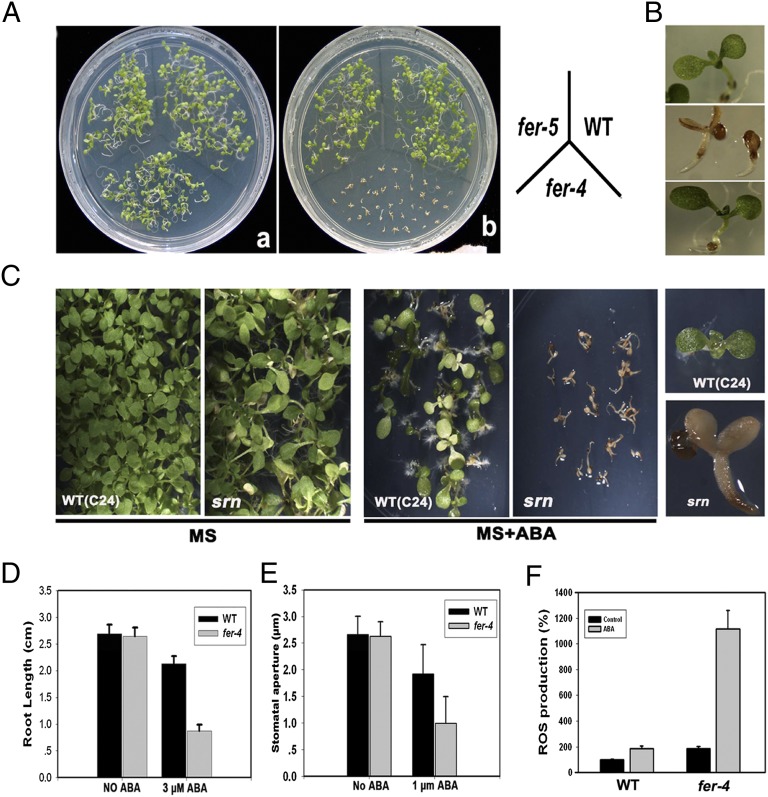
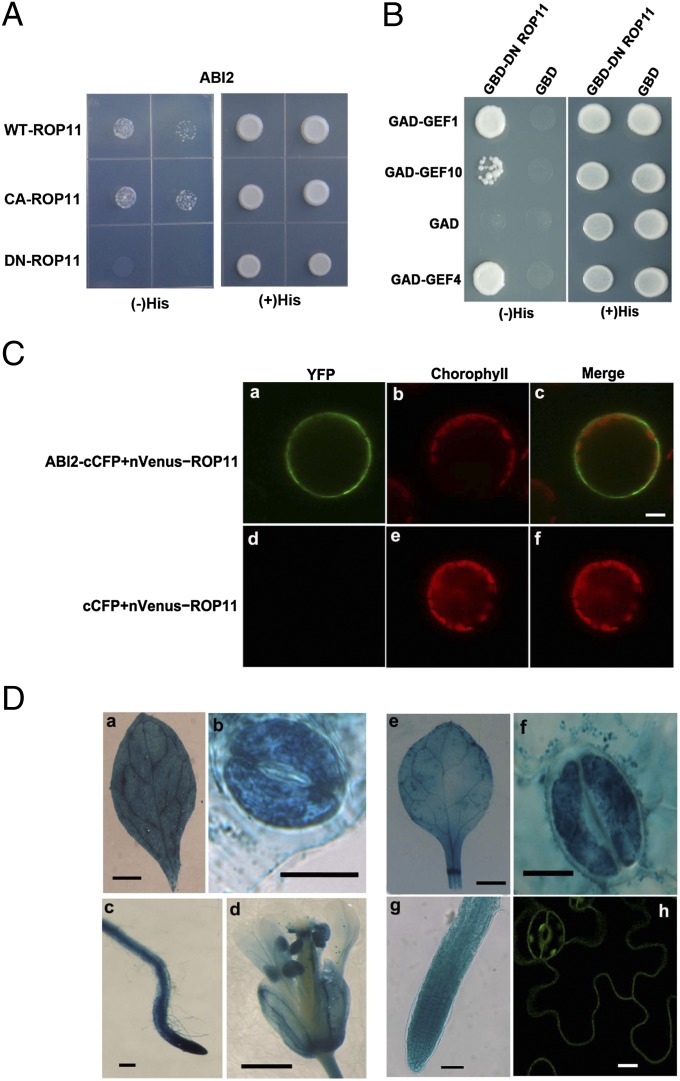
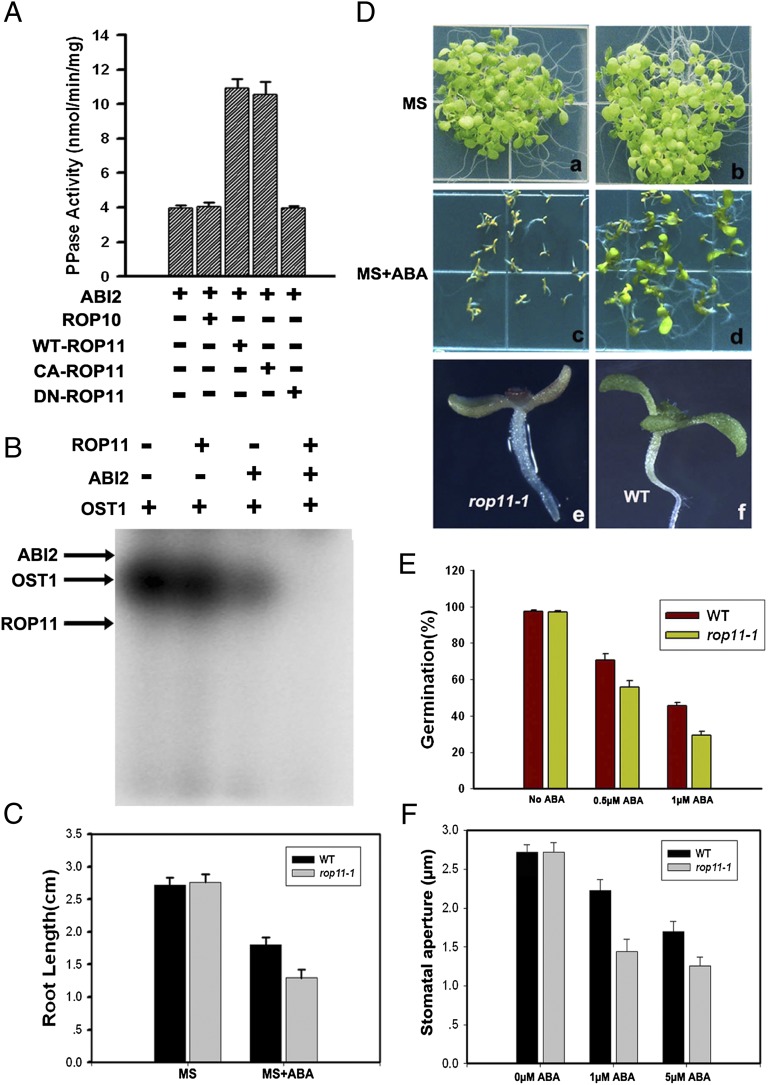
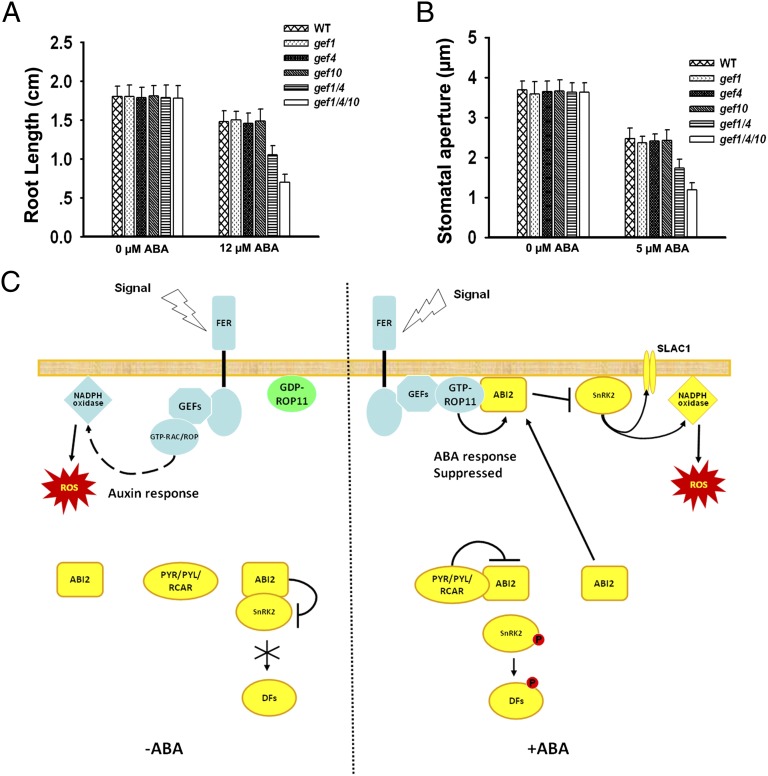
Plant growth and development are controlled by a delicate balance of hormonal cues. Growth-promoting hormones and growth-inhibiting counterparts often antagonize each other in their action, but the molecular mechanisms underlying these events remain largely unknown. Here, we report a cross-talk mechanism that enables a receptor-like kinase, FERONIA (FER), a positive regulator of auxin-promoted growth, to suppress the abscisic acid (ABA) response through activation of ABI2, a negative regulator of ABA signaling. The FER pathway consists of a FER kinase interacting with guanine exchange factors GEF1, GEF4, and GEF10 that, in turn, activate GTPase ROP11/ARAC10. Arabidopsis mutants disrupted in any step of the FER pathway, including fer, gef1gef4gef10, or rop11/arac10, all displayed an ABA-hypersensitive response, implicating the FER pathway in the suppression mechanism. In search of the target for the FER pathway, we found that the ROP11/ARAC10 protein physically interacted with the ABI2 phosphatase and enhanced its activity, thereby linking the FER pathway with the inhibition of ABA signaling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
FERONIA interacts with ABI2-type phosphatases to facilitate signaling cross-talk between abscisic acid and RALF peptide in Arabidopsis.Proc Natl Acad Sci U S A. 2016 Sep 13;113(37):E5519-27. doi: 10.1073/pnas.1608449113. Epub 2016 Aug 26. Proc Natl Acad Sci U S A. 2016. PMID: 27566404 Free PMC article.
-
FERONIA receptor-like kinase regulates RHO GTPase signaling of root hair development.Proc Natl Acad Sci U S A. 2010 Oct 12;107(41):17821-6. doi: 10.1073/pnas.1005366107. Epub 2010 Sep 27. Proc Natl Acad Sci U S A. 2010. PMID: 20876100 Free PMC article.
-
Glycosylphosphatidylinositol-anchored proteins as chaperones and co-receptors for FERONIA receptor kinase signaling in Arabidopsis.Elife. 2015 Jun 8;4:e06587. doi: 10.7554/eLife.06587. Elife. 2015. PMID: 26052747 Free PMC article.
-
FERONIA Receptor Kinase at the Crossroads of Hormone Signaling and Stress Responses.Plant Cell Physiol. 2017 Jul 1;58(7):1143-1150. doi: 10.1093/pcp/pcx048. Plant Cell Physiol. 2017. PMID: 28444222 Review.
-
Jack of all trades: crosstalk between FERONIA signaling and hormone pathways.J Exp Bot. 2025 May 10;76(7):1907-1920. doi: 10.1093/jxb/eraf071. J Exp Bot. 2025. PMID: 39972666 Free PMC article. Review.
Cited by
-
The Arabidopsis small GTPase AtRAC7/ROP9 is a modulator of auxin and abscisic acid signalling.J Exp Bot. 2013 Aug;64(11):3425-37. doi: 10.1093/jxb/ert179. J Exp Bot. 2013. PMID: 23918972 Free PMC article.
-
Genome-wide association study revealed genetic variations of ABA sensitivity controlled by multiple stress-related genes in rice.Stress Biol. 2021 Oct 7;1(1):10. doi: 10.1007/s44154-021-00011-4. Stress Biol. 2021. PMID: 37676585 Free PMC article.
-
OsMADS23 phosphorylated by SAPK9 confers drought and salt tolerance by regulating ABA biosynthesis in rice.PLoS Genet. 2021 Aug 3;17(8):e1009699. doi: 10.1371/journal.pgen.1009699. eCollection 2021 Aug. PLoS Genet. 2021. PMID: 34343171 Free PMC article.
-
Genome-wide identification and analysis of Catharanthus roseus RLK1-like kinases in rice.Planta. 2015 Mar;241(3):603-13. doi: 10.1007/s00425-014-2203-2. Epub 2014 Nov 16. Planta. 2015. PMID: 25399351
-
RALF-FERONIA Signaling: Linking Plant Immune Response with Cell Growth.Plant Commun. 2020 Jun 11;1(4):100084. doi: 10.1016/j.xplc.2020.100084. eCollection 2020 Jul 13. Plant Commun. 2020. PMID: 33367248 Free PMC article. Review.
References
-
- Bellandi DM, Dorffling K. Effect of abscisic acid and other plant hormones on growth of apical and lateral buds of seedlings. Physiol Plant. 1974;32:369–372.
-
- Pilet PE. Differential growth and hormone redistribution in gravireacting maize roots. Environ Exp Bot. 1989;29:37–45. - PubMed
-
- Raghavendra AS, Gonugunta VK, Christmann A, Grill E. ABA perception and signalling. Trends Plant Sci. 2010;15:395–401. - PubMed
-
- Cutler SR, Rodriguez PL, Finkelstein RR, Abrams SR. Abscisic acid: Emergence of a core signaling network. Annu Rev Plant Biol. 2010;61:651–679. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
