

Eilat virus, a unique alphavirus with host range restricted to insects by RNA replication
- PMID: 22908261
- PMCID: PMC3437828
- DOI: 10.1073/pnas.1204787109
Eilat virus, a unique alphavirus with host range restricted to insects by RNA replication
Abstract
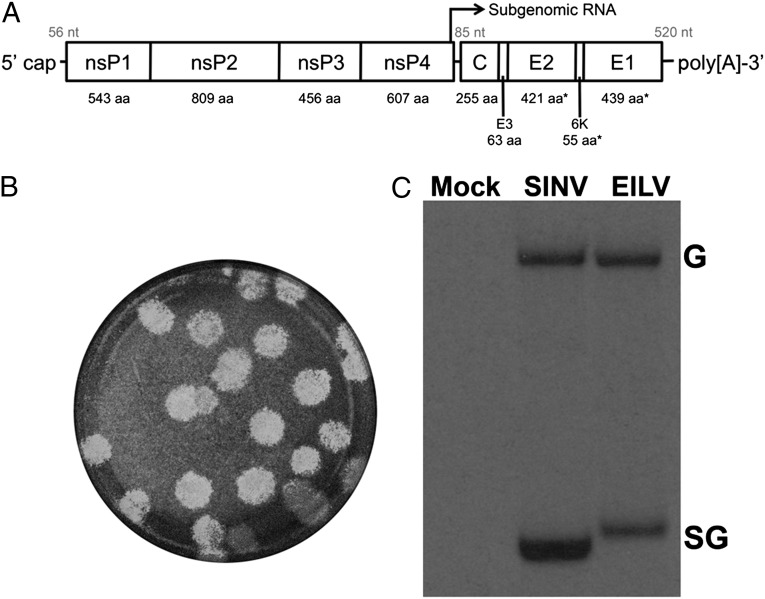
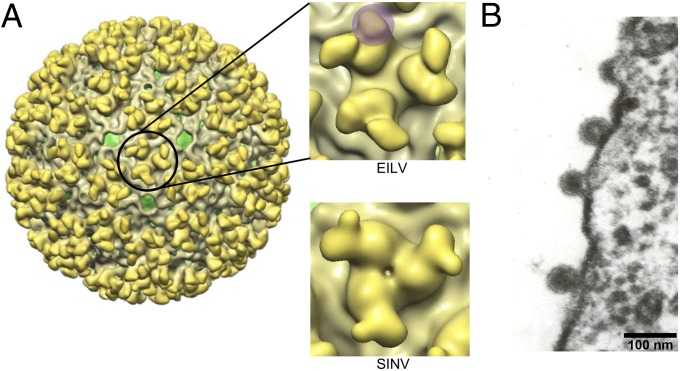
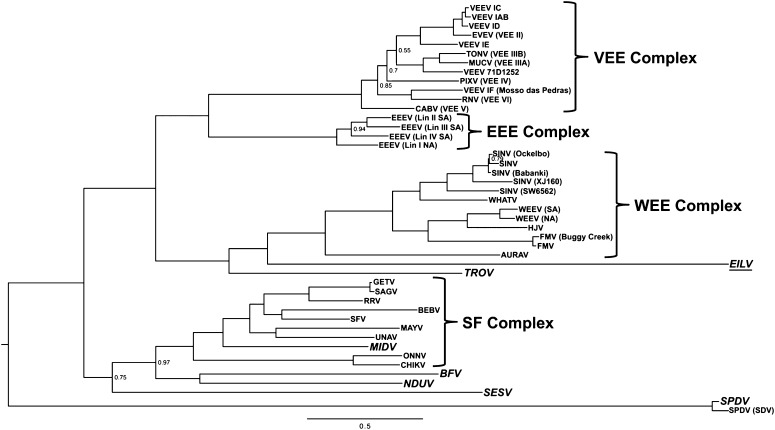
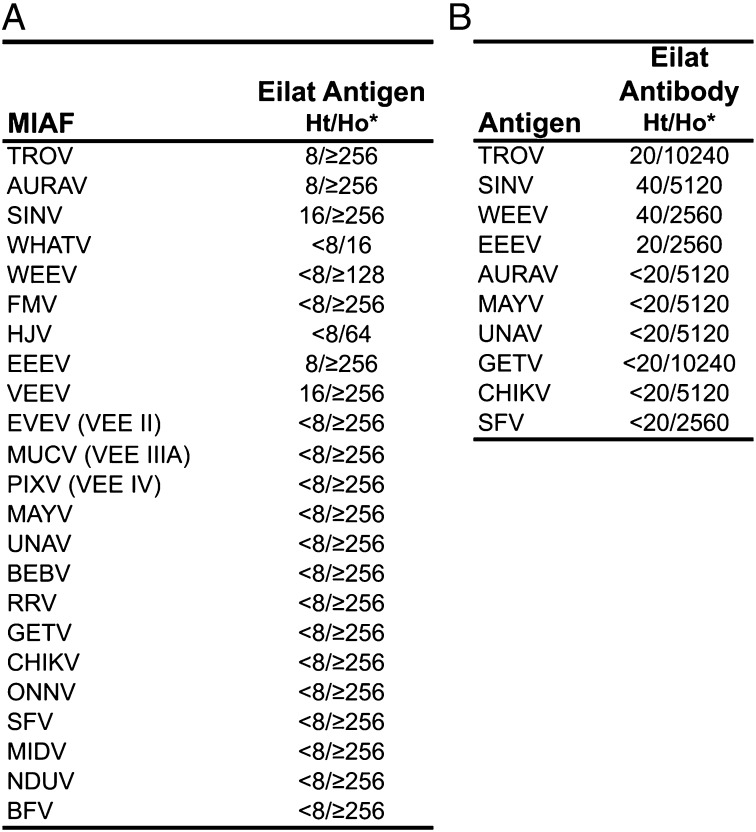
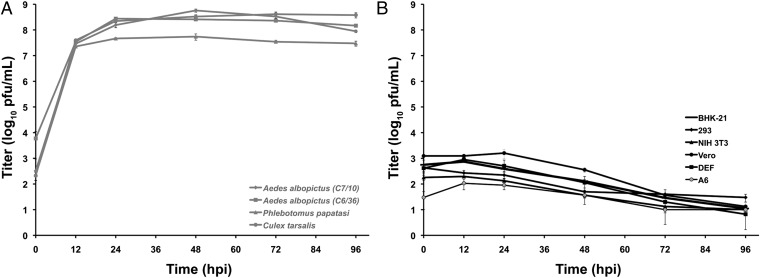
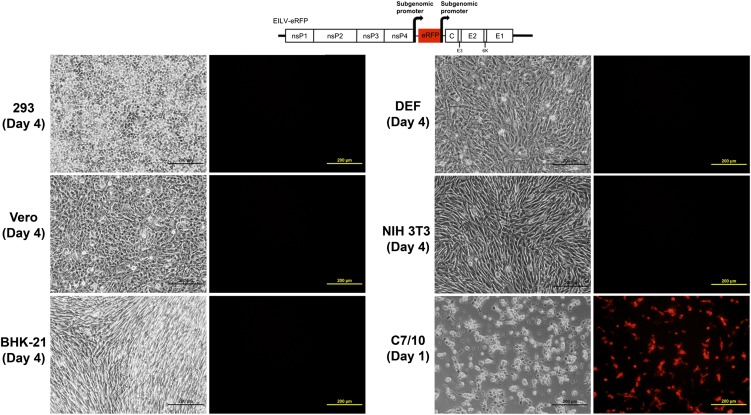
Most alphaviruses and many other arboviruses are mosquito-borne and exhibit a broad host range, infecting many different vertebrates including birds, rodents, equids, humans, and nonhuman primates. Consequently, they can be propagated in most vertebrate and insect cell cultures. This ability of arboviruses to infect arthropods and vertebrates is usually essential for their maintenance in nature. However, several flaviviruses have recently been described that infect mosquitoes but not vertebrates, although the mechanism of their host restriction has not been determined. Here we describe a unique alphavirus, Eilat virus (EILV), isolated from a pool of Anopheles coustani mosquitoes from the Negev desert of Israel. Phylogenetic analyses placed EILV as a sister to the Western equine encephalitis antigenic complex within the main clade of mosquito-borne alphaviruses. Electron microscopy revealed that, like other alphaviruses, EILV virions were spherical, 70 nm in diameter, and budded from the plasma membrane of mosquito cells in culture. EILV readily infected a variety of insect cells with little overt cytopathic effect. However, in contrast to typical mosquito-borne alphaviruses, EILV could not infect mammalian or avian cell lines, and viral as well as RNA replication could not be detected at 37 °C or 28 °C. Evolutionarily, these findings suggest that EILV lost its ability to infect vertebrate cells. Thus, EILV seems to be mosquito-specific and represents a previously undescribed complex within the genus Alphavirus. Reverse genetic studies of EILV may facilitate the discovery of determinants of alphavirus host range that mediate disease emergence.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kuhn RJ. Togaviridae: The viruses and their replication. In: Knipe DM, Howley PM, editors. Fields Virology. 5th ed. Baltimore: Lippincott Williams and Wilkins; 2007. pp. 1001–1022.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
