

Function of alternative splicing
- PMID: 22909801
- PMCID: PMC5632952
- DOI: 10.1016/j.gene.2012.07.083
Function of alternative splicing
Abstract
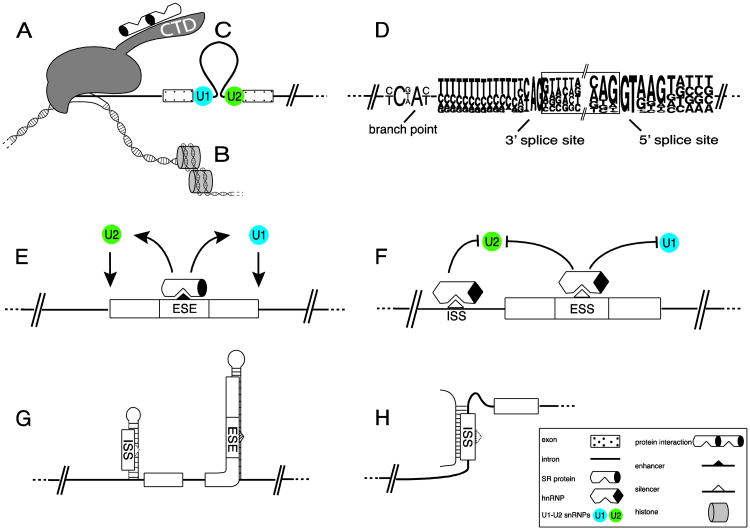
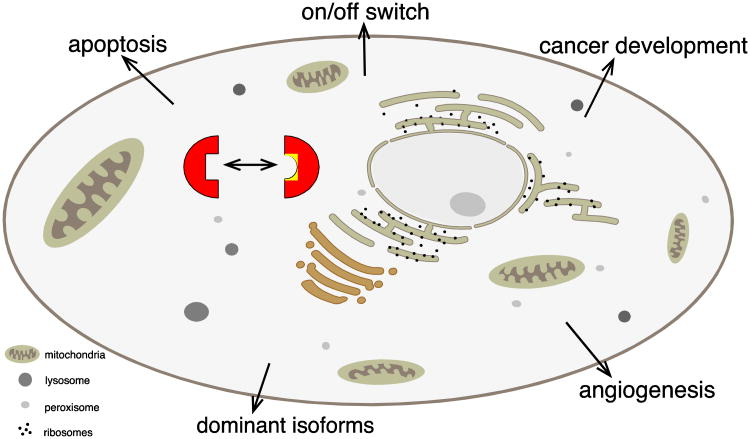
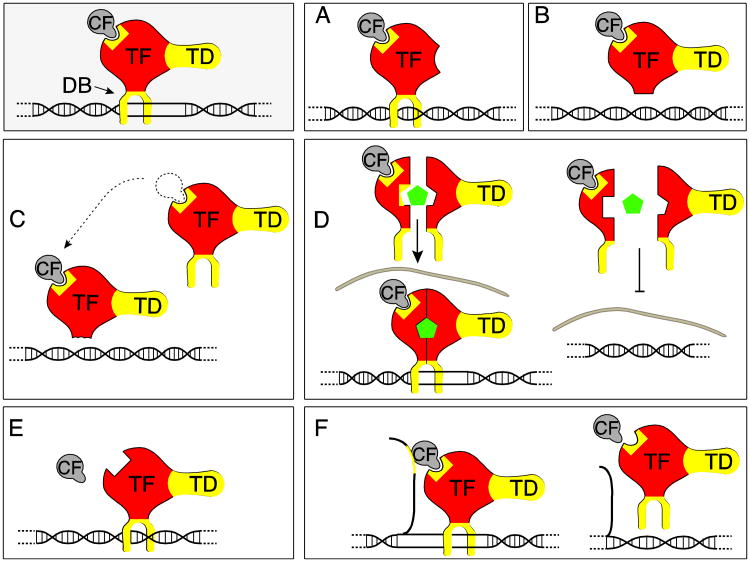
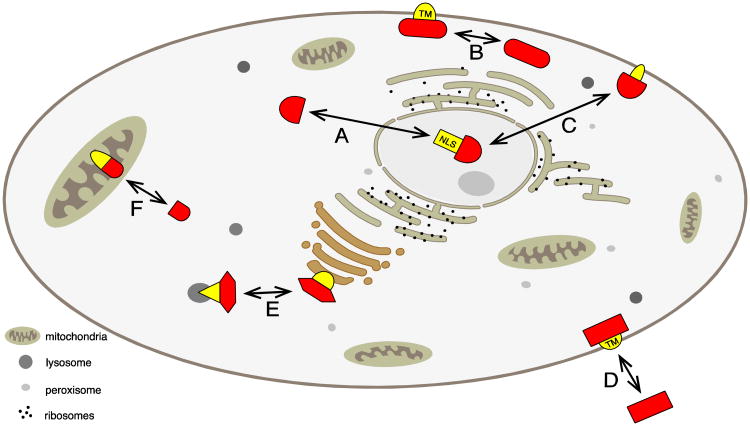
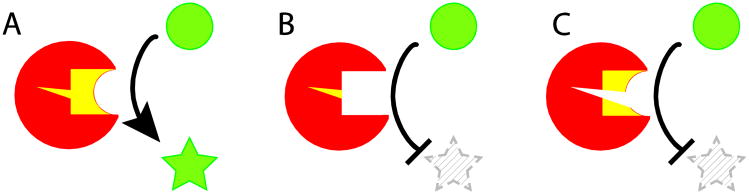
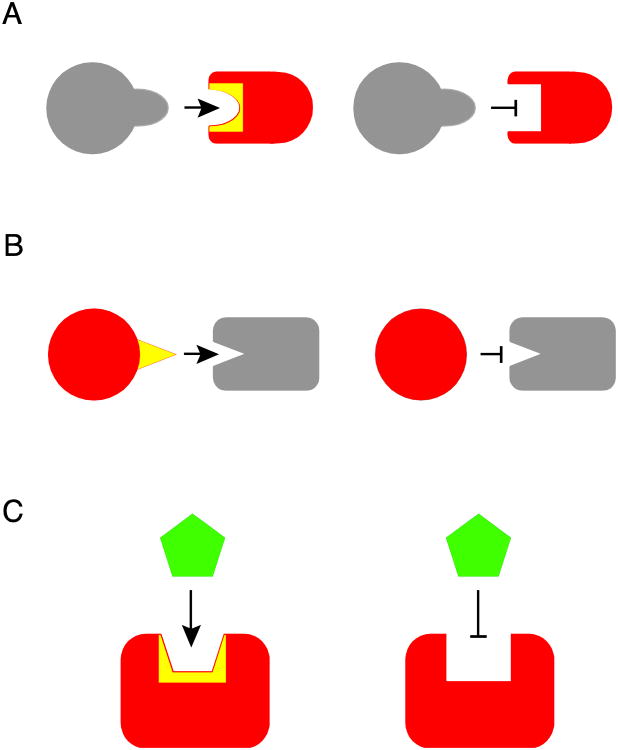
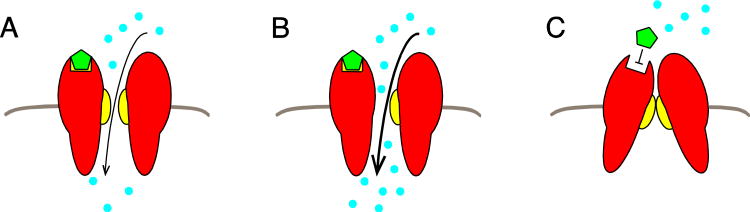
Almost all polymerase II transcripts undergo alternative pre-mRNA splicing. Here, we review the functions of alternative splicing events that have been experimentally determined. The overall function of alternative splicing is to increase the diversity of mRNAs expressed from the genome. Alternative splicing changes proteins encoded by mRNAs, which has profound functional effects. Experimental analysis of these protein isoforms showed that alternative splicing regulates binding between proteins, between proteins and nucleic acids as well as between proteins and membranes. Alternative splicing regulates the localization of proteins, their enzymatic properties and their interaction with ligands. In most cases, changes caused by individual splicing isoforms are small. However, cells typically coordinate numerous changes in 'splicing programs', which can have strong effects on cell proliferation, cell survival and properties of the nervous system. Due to its widespread usage and molecular versatility, alternative splicing emerges as a central element in gene regulation that interferes with almost every biological function analyzed.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
References
-
- Adams PJ, Garcia E, David LS, Mulatz KJ, Spacey SD, Snutch TP. Ca(V)2.1 P/Q-type calcium channel alternative splicing affects the functional impact of familial hemiplegic migraine mutations: implications for calcium channelopathies. Channels (Austin) 2009;3:110–21. - PubMed
-
- Adato A, Lefevre G, Delprat B, Michel V, Michalski N, Chardenoux S, Weil D, El-Amraoui A, Petit C. Usherin, the defective protein in Usher syndrome type IIA, is likely to be a component of interstereocilia ankle links in the inner ear sensory cells. Human molecular genetics. 2005;14:3921–32. - PubMed
-
- Ahn J, Febbraio M, Silverstein RL. A novel isoform of human Golgi complex-localized glycoprotein-1 (also known as E-selectin ligand-1, MG-160 and cysteine-rich fibroblast growth factor receptor) targets differential subcellular localization. J Cell Sci. 2005;118:1725–31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
