

Analysis of histone chaperone antisilencing function 1 interactions
- PMID: 22910209
- PMCID: PMC3493145
- DOI: 10.1016/B978-0-12-391940-3.00010-X
Analysis of histone chaperone antisilencing function 1 interactions
Abstract
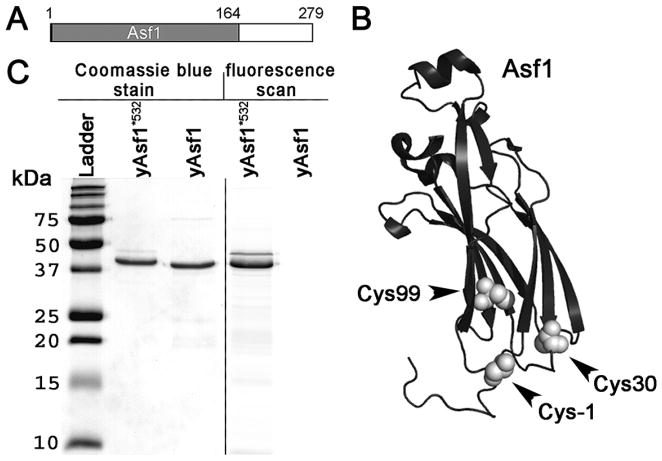
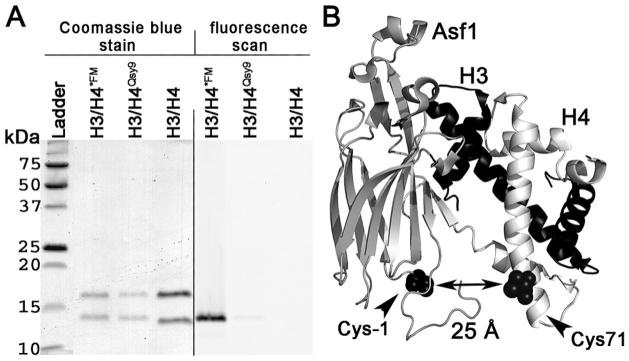
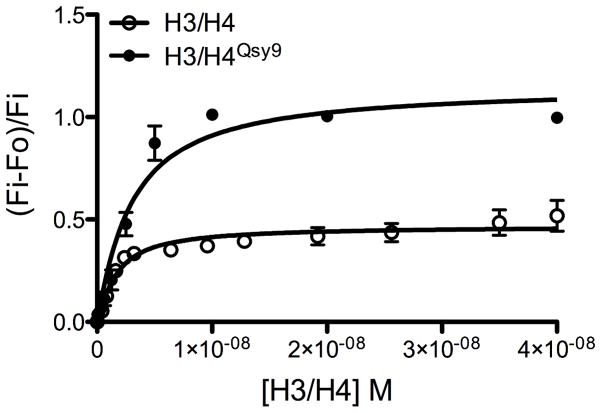
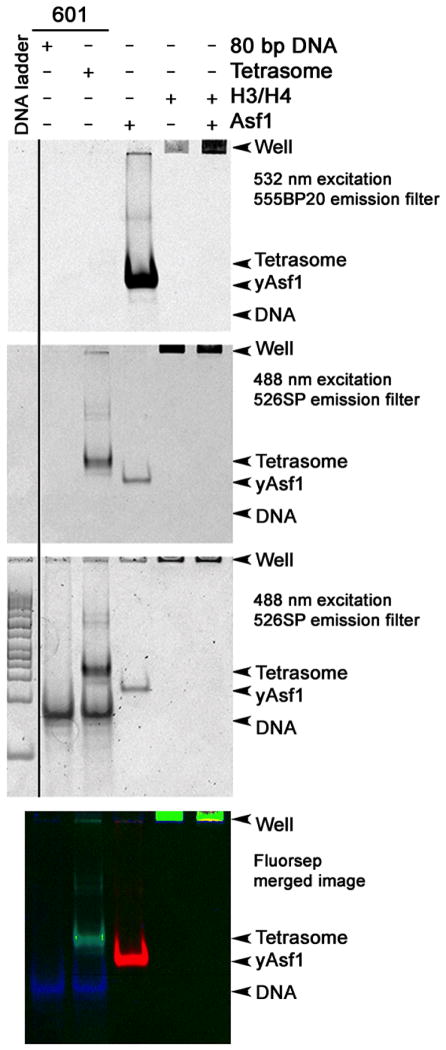
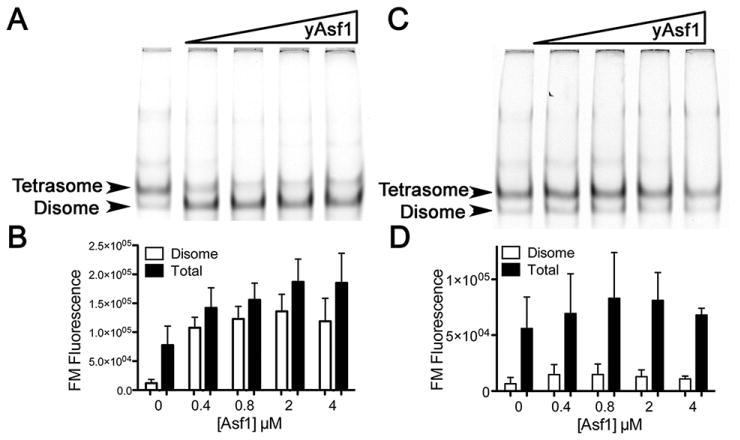
The assembly and disassembly of chromatin impacts all DNA-dependent processes in eukaryotes. These processes are intricately regulated through stepwise mechanisms, requiring multiple proteins, posttranslational modifications, and remodeling enzymes, as well as specific proteins to chaperone the highly basic and aggregation-prone histone proteins. The histone chaperones are acidic proteins that perform the latter function by maintaining the stability of the histones when they are not associated with DNA and guiding the deposition and removal of histones from DNA. Understanding the thermodynamics of these processes provides deeper insights into the mechanisms of chromatin assembly and disassembly. Here we describe complementary thermodynamic and biochemical approaches for analysis of the interactions of a major chaperone of the H3/H4 dimer, anti-silencing function 1 (Asf1) with histones H3/H4, and DNA. Fluorescence quenching approaches are useful for measuring the binding affinity of Asf1 for histones H3/H4 under equilibrium conditions. Electrophoretic mobility shift analyses are useful for examining Asf1-mediated tetrasome (H3/H4-DNA) assembly and disassembly processes. These approaches potentially can be used more generally for the study of other histone chaperone-histone interactions and provide a means to dissect the role of posttranslational modifications and other factors that participate in chromatin dynamics.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
The activity of the histone chaperone yeast Asf1 in the assembly and disassembly of histone H3/H4-DNA complexes.Nucleic Acids Res. 2011 Jul;39(13):5449-58. doi: 10.1093/nar/gkr097. Epub 2011 Mar 29. Nucleic Acids Res. 2011. PMID: 21447559 Free PMC article.
-
Structure and function of the histone chaperone CIA/ASF1 complexed with histones H3 and H4.Nature. 2007 Mar 15;446(7133):338-41. doi: 10.1038/nature05613. Epub 2007 Feb 11. Nature. 2007. PMID: 17293877
-
Structural basis for the histone chaperone activity of Asf1.Cell. 2006 Nov 3;127(3):495-508. doi: 10.1016/j.cell.2006.08.047. Cell. 2006. PMID: 17081973 Free PMC article.
-
The histone chaperone Asf1 at the crossroads of chromatin and DNA checkpoint pathways.Chromosoma. 2007 Apr;116(2):79-93. doi: 10.1007/s00412-006-0087-z. Epub 2006 Dec 19. Chromosoma. 2007. PMID: 17180700 Review.
-
Histone transfer among chaperones.Biochem Soc Trans. 2012 Apr;40(2):357-63. doi: 10.1042/BST20110737. Biochem Soc Trans. 2012. PMID: 22435812 Free PMC article. Review.
Cited by
-
The C terminus of the histone chaperone Asf1 cross-links to histone H3 in yeast and promotes interaction with histones H3 and H4.Mol Cell Biol. 2013 Feb;33(3):605-21. doi: 10.1128/MCB.01053-12. Epub 2012 Nov 26. Mol Cell Biol. 2013. PMID: 23184661 Free PMC article.
-
sNASP and ASF1A function through both competitive and compatible modes of histone binding.Nucleic Acids Res. 2017 Jan 25;45(2):643-656. doi: 10.1093/nar/gkw892. Epub 2016 Oct 5. Nucleic Acids Res. 2017. PMID: 28123037 Free PMC article.
-
Abo1 ATPase facilitates the dissociation of FACT from chromatin.Nucleic Acids Res. 2025 Feb 8;53(4):gkae1229. doi: 10.1093/nar/gkae1229. Nucleic Acids Res. 2025. PMID: 39676666 Free PMC article.
-
Site specificity analysis of Piccolo NuA4-mediated acetylation for different histone complexes.Biochem J. 2015 Dec 1;472(2):239-48. doi: 10.1042/BJ20150654. Epub 2015 Sep 29. Biochem J. 2015. PMID: 26420880 Free PMC article.
-
CAF-1-induced oligomerization of histones H3/H4 and mutually exclusive interactions with Asf1 guide H3/H4 transitions among histone chaperones and DNA.Nucleic Acids Res. 2012 Dec;40(22):11229-39. doi: 10.1093/nar/gks906. Epub 2012 Oct 2. Nucleic Acids Res. 2012. PMID: 23034810 Free PMC article.
References
-
- Akey CW, Luger K. Histone chaperones and nucleosome assembly. Curr Opin Struct Biol. 2003;13(1):6–14. - PubMed
-
- Alilat M, Sivolob A, Revet B, Prunell A. Nucleosome dynamics. Protein and DNA contributions in the chiral transition of the tetrasome, the histone (H3–H4)2 tetramer-DNA particle. J Mol Biol. 1999;291(4):815–41. - PubMed
-
- Amin AD, Vishnoi N, Prochasson P. A global requirement for the HIR complex in the assembly of chromatin’. Biochim Biophys Acta 2011 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
