

Non-transgenic genome modifications in a hemimetabolous insect using zinc-finger and TAL effector nucleases
- PMID: 22910363
- PMCID: PMC3432469
- DOI: 10.1038/ncomms2020
Non-transgenic genome modifications in a hemimetabolous insect using zinc-finger and TAL effector nucleases
Abstract
Hemimetabolous, or incompletely metamorphosing, insects are phylogenetically relatively basal and comprise many pests. However, the absence of a sophisticated genetic model system, or targeted gene-manipulation system, has limited research on hemimetabolous species. Here we use zinc-finger nuclease and transcription activator-like effector nuclease technologies to produce genetic knockouts in the hemimetabolous insect Gryllus bimaculatus. Following the microinjection of mRNAs encoding zinc-finger nucleases or transcription activator-like effector nucleases into cricket embryos, targeting of a transgene or endogenous gene results in sequence-specific mutations. Up to 48% of founder animals transmit disrupted gene alleles after zinc-finger nucleases microinjection compared with 17% after microinjection of transcription activator-like effector nucleases. Heterozygous offspring is selected using mutation detection assays that use a Surveyor (Cel-I) nuclease, and subsequent sibling crosses create homozygous knockout crickets. This approach is independent from a mutant phenotype or the genetic tractability of the organism of interest and can potentially be applied to manage insect pests using a non-transgenic strategy.
Figures
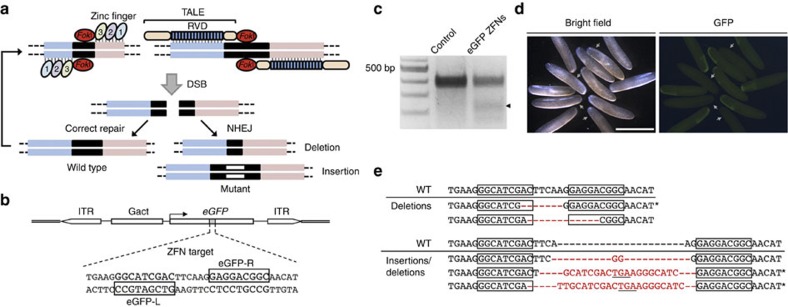
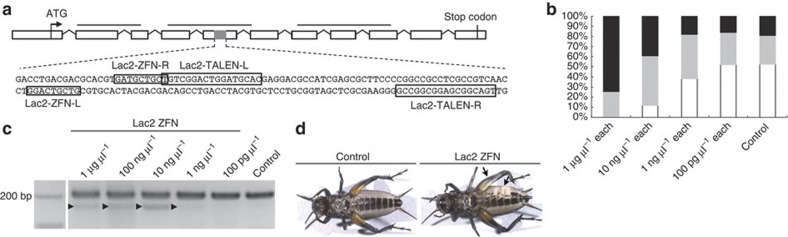

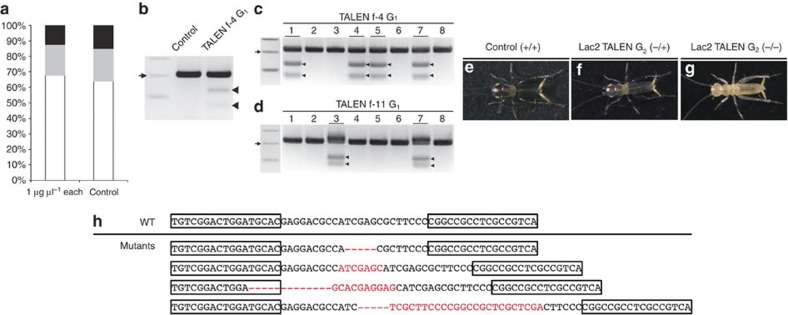
References
-
- Rothstein R. J. One-step gene disruption in yeast. Methods Enzymol. 101, 202–211 (1983). - PubMed
-
- Rong Y. S. & Golic K. G. in Insect Transgenesis Methods and Applications (eds Handler, A. M., James, A. A.) 53–75 CRC Press, 2000.
-
- Capecchi M. R. Gene targeting in mice: functional analysis of the mammalian genome for the twenty-first century. Nat. Rev. Genet. 6, 507–512 (2005). - PubMed
-
- Porteus M. H. & Carroll D. Gene targeting using zinc finger nucleases. Nat. Biotechnol. 23, 967–973 (2005). - PubMed
-
- Miller J. C. et al.. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 29, 143–148 (2011). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
