

Pregnancy-induced noncoding RNA (PINC) associates with polycomb repressive complex 2 and regulates mammary epithelial differentiation
- PMID: 22911650
- PMCID: PMC3406180
- DOI: 10.1371/journal.pgen.1002840
Pregnancy-induced noncoding RNA (PINC) associates with polycomb repressive complex 2 and regulates mammary epithelial differentiation
Abstract
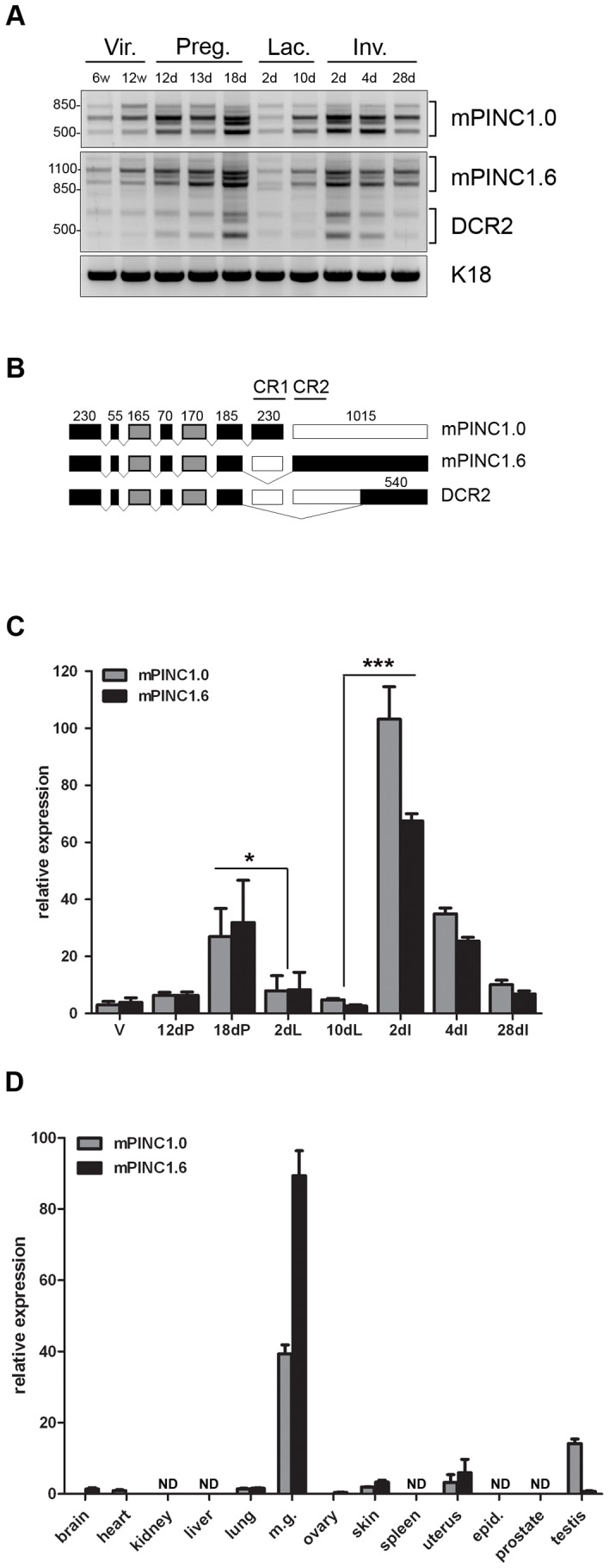
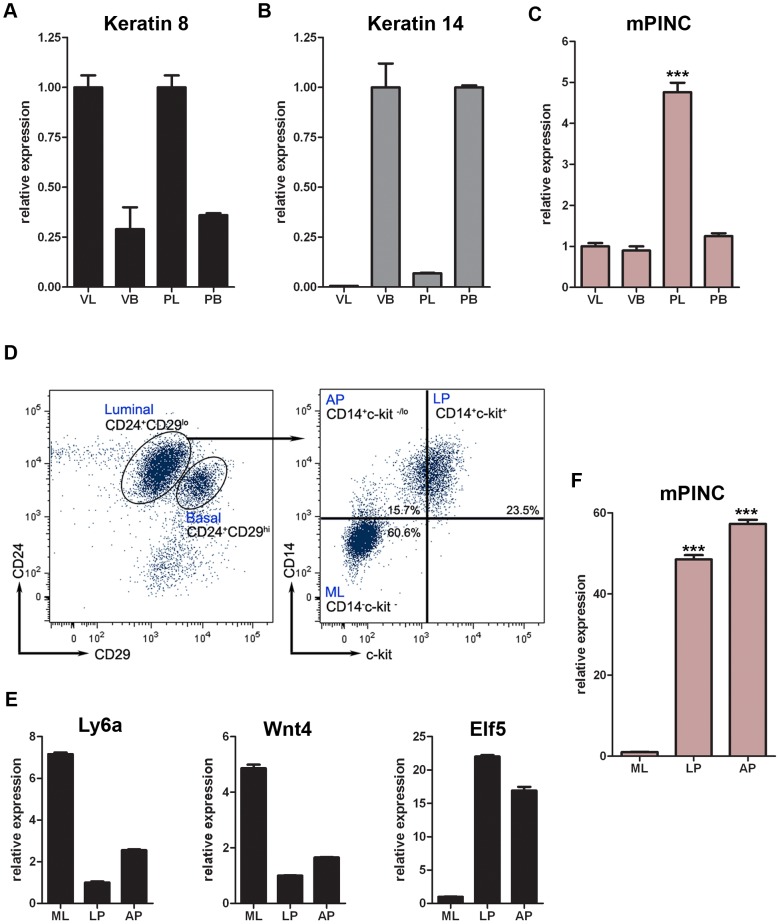
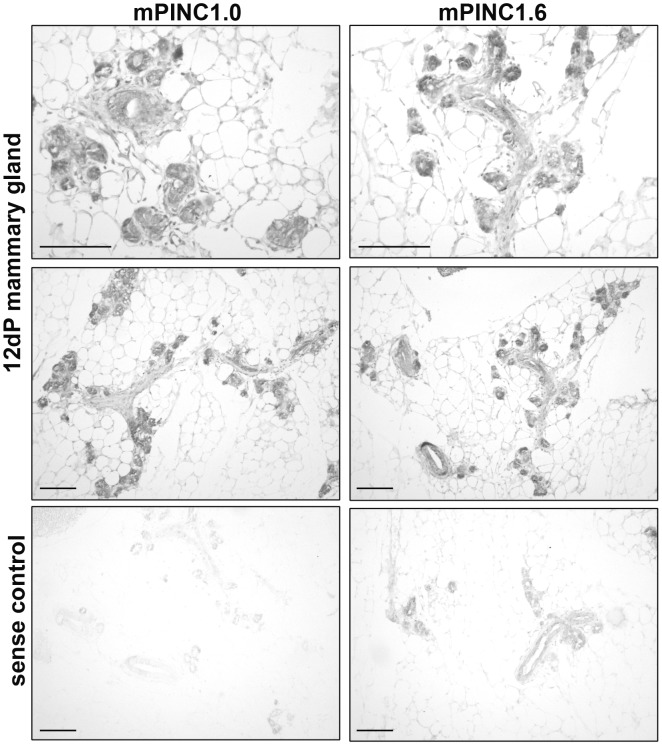
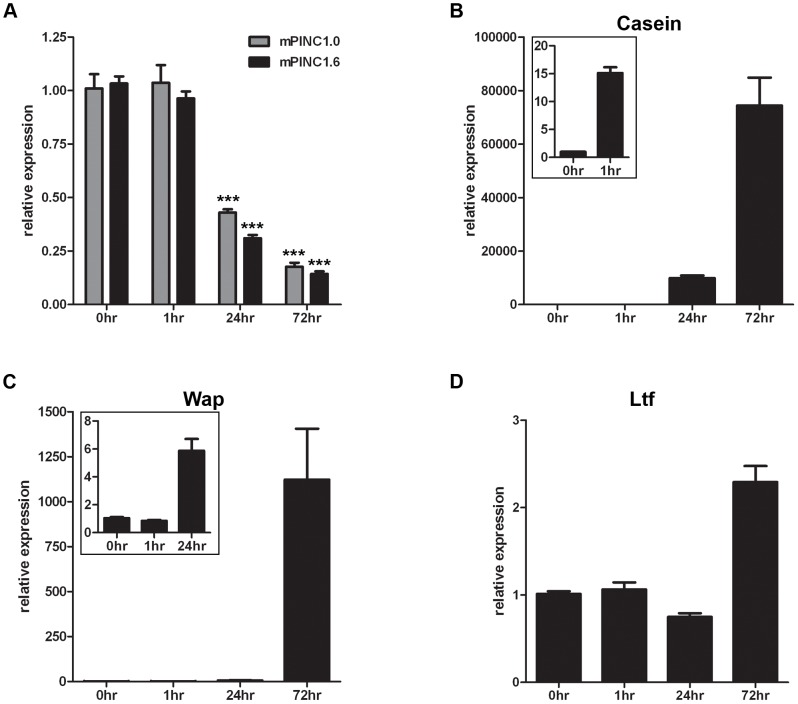
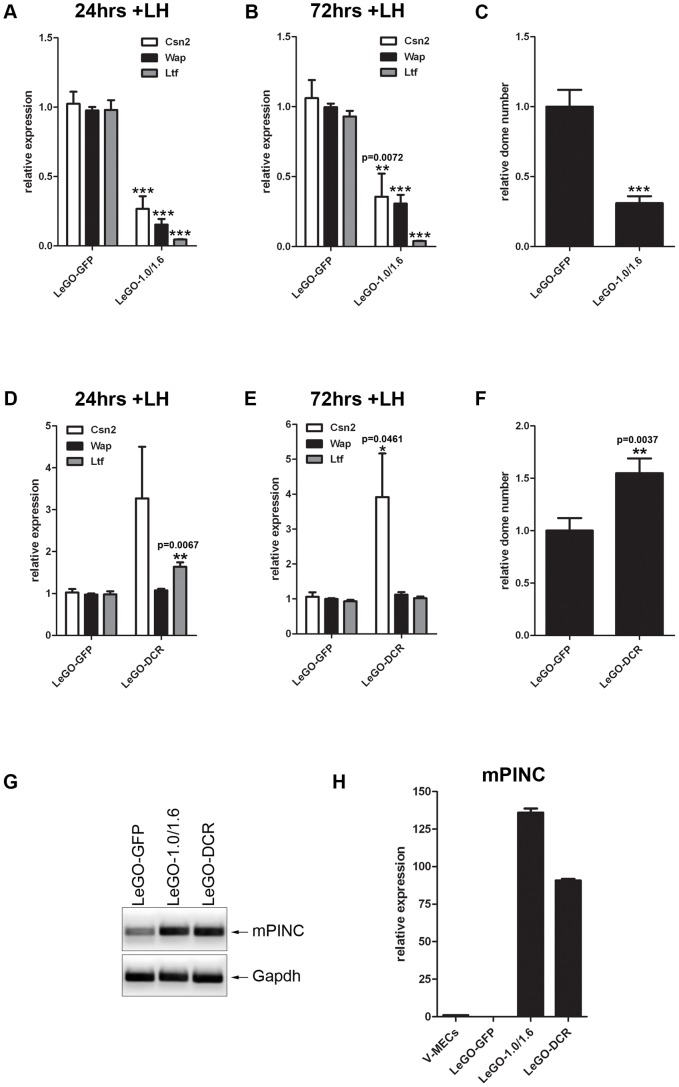
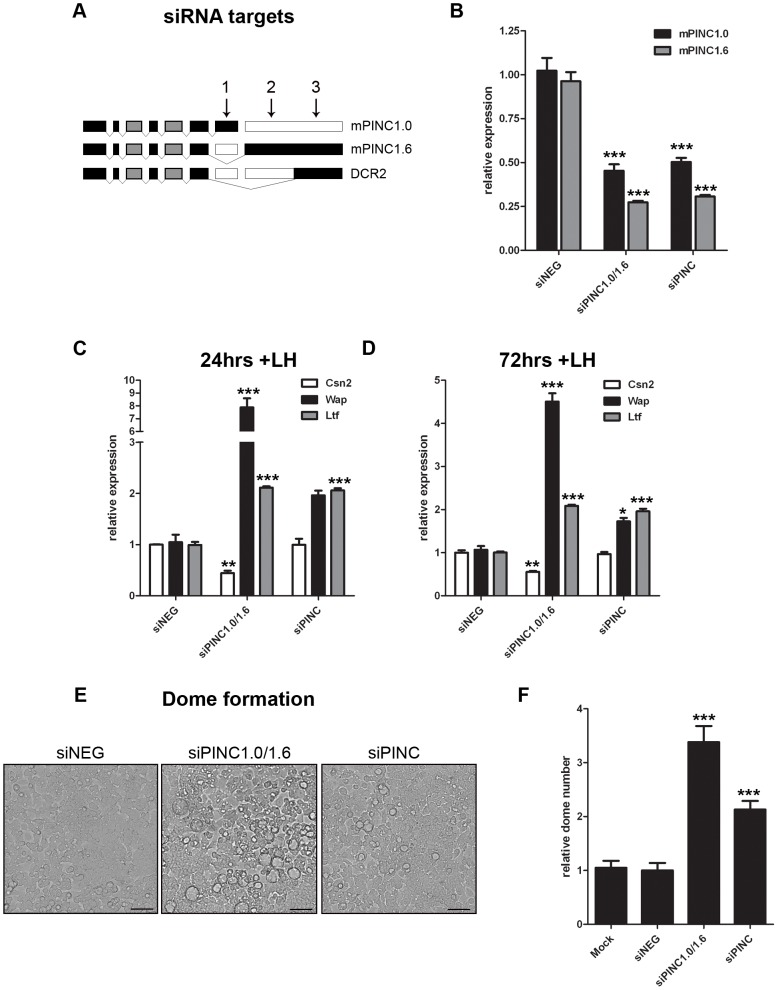
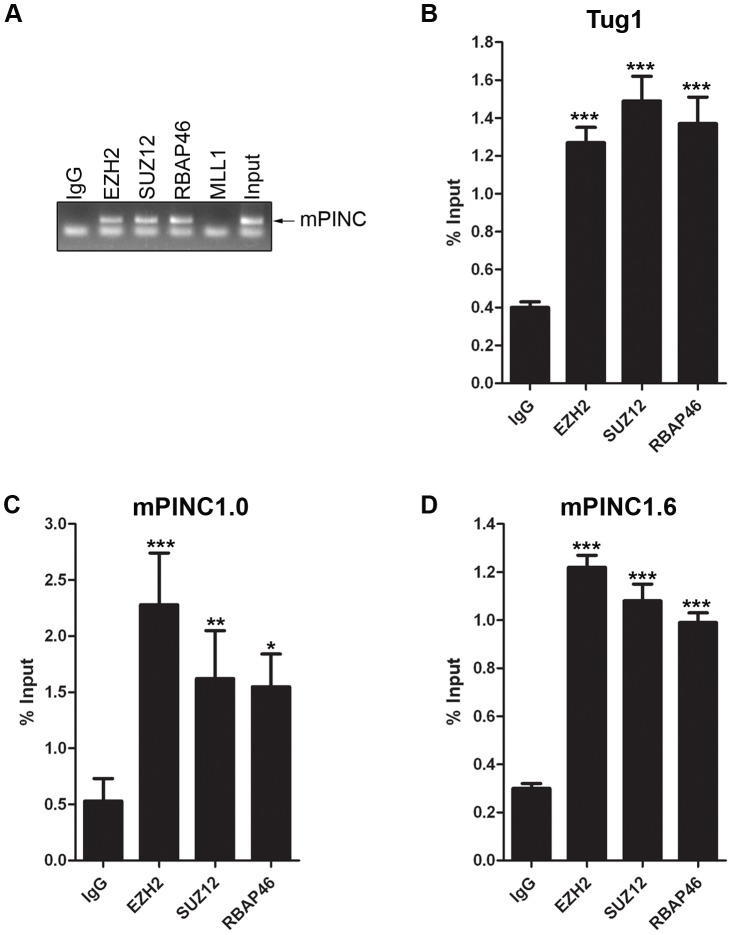
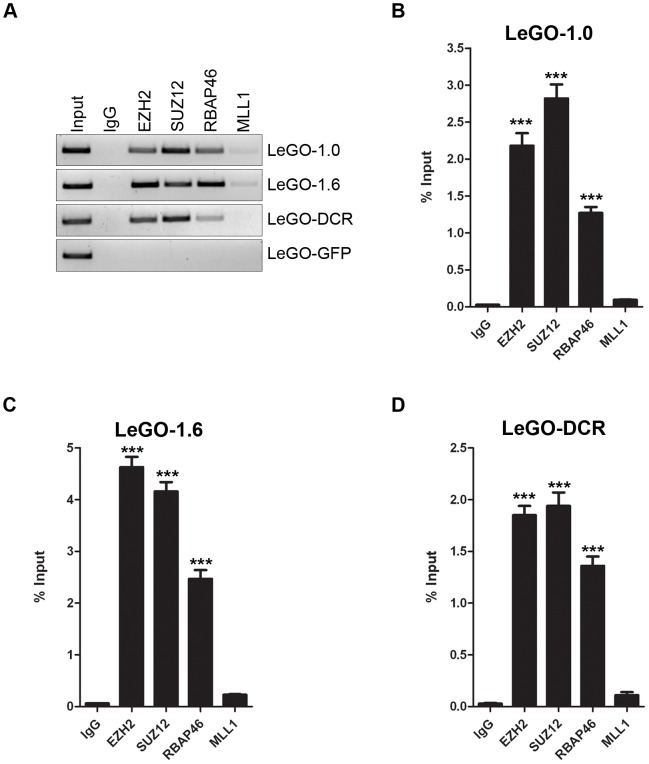
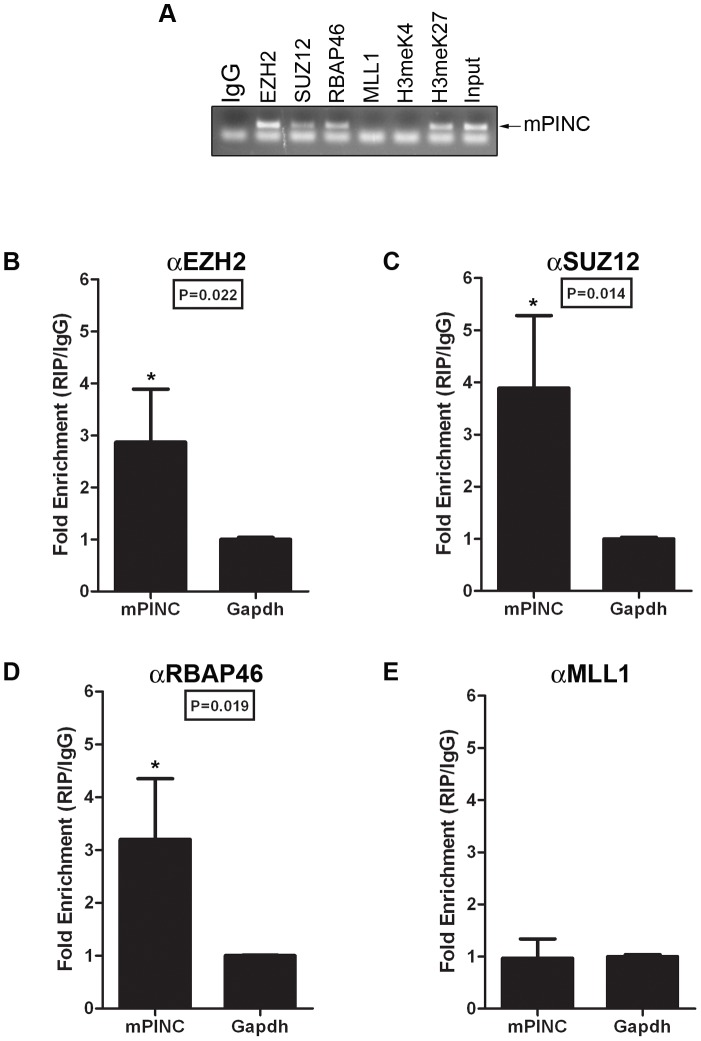
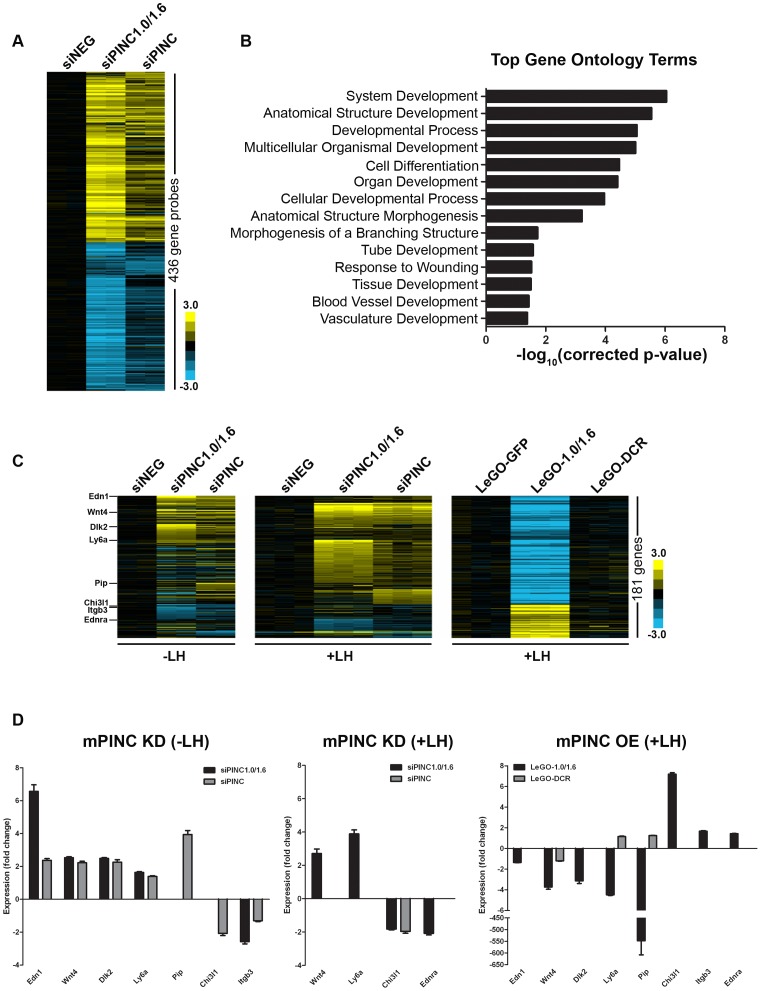
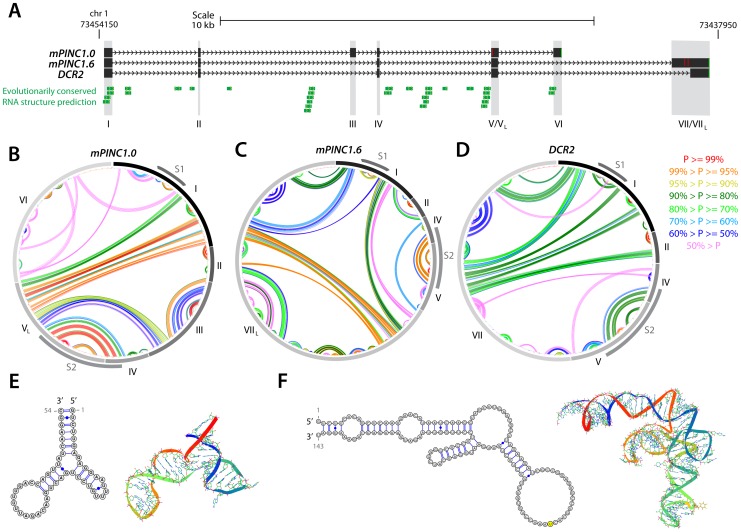
Pregnancy-induced noncoding RNA (PINC) and retinoblastoma-associated protein 46 (RbAp46) are upregulated in alveolar cells of the mammary gland during pregnancy and persist in alveolar cells that remain in the regressed lobules following involution. The cells that survive involution are thought to function as alveolar progenitor cells that rapidly differentiate into milk-producing cells in subsequent pregnancies, but it is unknown whether PINC and RbAp46 are involved in maintaining this progenitor population. Here, we show that, in the post-pubertal mouse mammary gland, mPINC is enriched in luminal and alveolar progenitors. mPINC levels increase throughout pregnancy and then decline in early lactation, when alveolar cells undergo terminal differentiation. Accordingly, mPINC expression is significantly decreased when HC11 mammary epithelial cells are induced to differentiate and produce milk proteins. This reduction in mPINC levels may be necessary for lactation, as overexpression of mPINC in HC11 cells blocks lactogenic differentiation, while knockdown of mPINC enhances differentiation. Finally, we demonstrate that mPINC interacts with RbAp46, as well as other members of the polycomb repressive complex 2 (PRC2), and identify potential targets of mPINC that are differentially expressed following modulation of mPINC expression levels. Taken together, our data suggest that mPINC inhibits terminal differentiation of alveolar cells during pregnancy to prevent abundant milk production and secretion until parturition. Additionally, a PRC2 complex that includes mPINC and RbAp46 may confer epigenetic modifications that maintain a population of mammary epithelial cells committed to the alveolar fate in the involuted gland.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
