

Conserved tyrosine kinase promotes the import of silencing RNA into Caenorhabditis elegans cells
- PMID: 22912399
- PMCID: PMC3437824
- DOI: 10.1073/pnas.1201153109
Conserved tyrosine kinase promotes the import of silencing RNA into Caenorhabditis elegans cells
Abstract
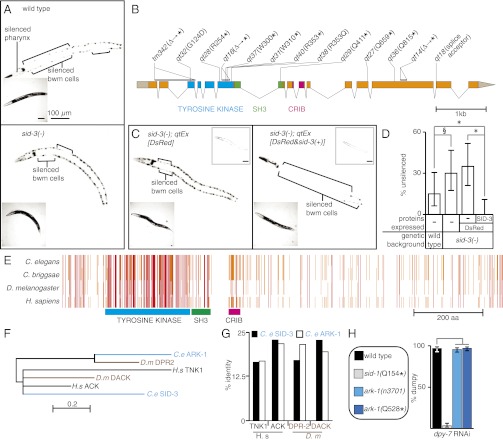
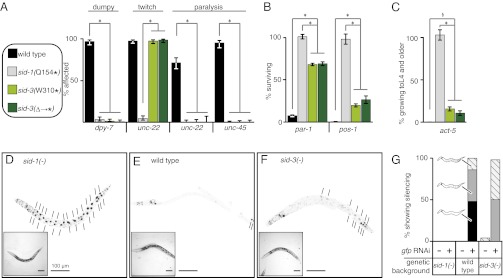
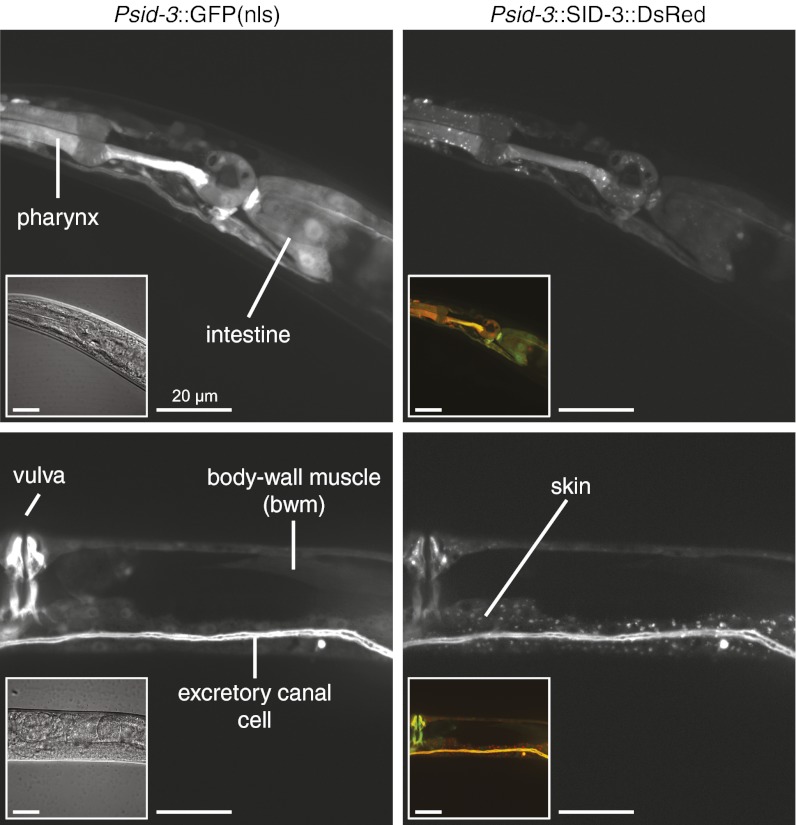
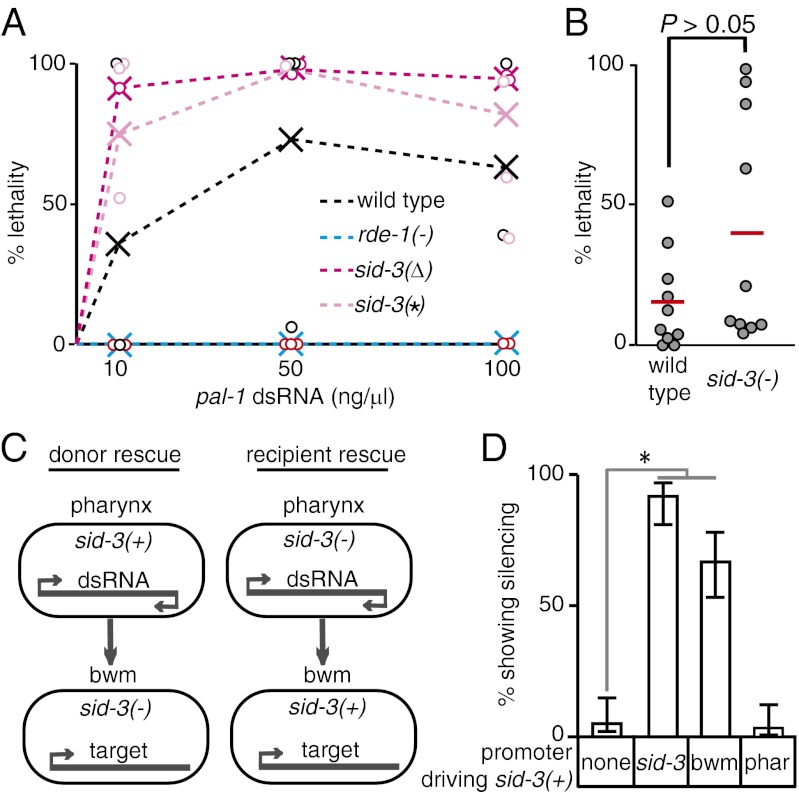
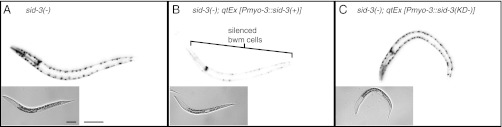
RNA silencing in Caenorhabditis elegans is transmitted between cells by the transport of double-stranded RNA (dsRNA). The efficiency of such transmission, however, depends on both the cell type and the environment. Here, we identify systemic RNAi defective-3 (SID-3) as a conserved tyrosine kinase required for the efficient import of dsRNA. Without SID-3, cells perform RNA silencing well but import dsRNA poorly. Upon overexpression of SID-3, cells import dsRNA more efficiently than do wild-type cells and such efficient import of dsRNA requires an intact SID-3 kinase domain. The mammalian homolog of SID-3, activated cdc-42-associated kinase (ACK), acts in many signaling pathways that respond to environmental changes and is known to directly associate with endocytic vesicles, which have been implicated in dsRNA transport. Therefore, our results suggest that the SID-3/ACK tyrosine kinase acts as a regulator of RNA import into animal cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Winston WM, Molodowitch C, Hunter CP. Systemic RNAi in C. elegans requires the putative transmembrane protein SID-1. Science. 2002;295:2456–2459. - PubMed
-
- Wolfrum C, et al. Mechanisms and optimization of in vivo delivery of lipophilic siRNAs. Nat Biotechnol. 2007;25:1149–1157. - PubMed
-
- Duxbury MS, Ashley SW, Whang EE. RNA interference: A mammalian SID-1 homologue enhances siRNA uptake and gene silencing efficacy in human cells. Biochem Biophys Res Commun. 2005;331:459–463. - PubMed
-
- Valadi H, et al. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9:654–659. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
