

The Burkholderia bcpAIOB genes define unique classes of two-partner secretion and contact dependent growth inhibition systems
- PMID: 22912595
- PMCID: PMC3415462
- DOI: 10.1371/journal.pgen.1002877
The Burkholderia bcpAIOB genes define unique classes of two-partner secretion and contact dependent growth inhibition systems
Abstract
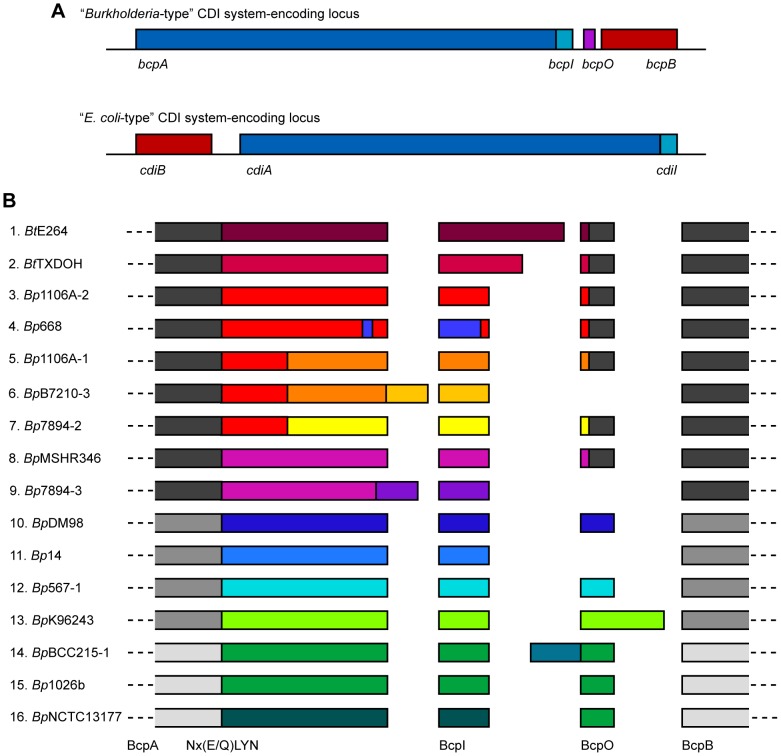
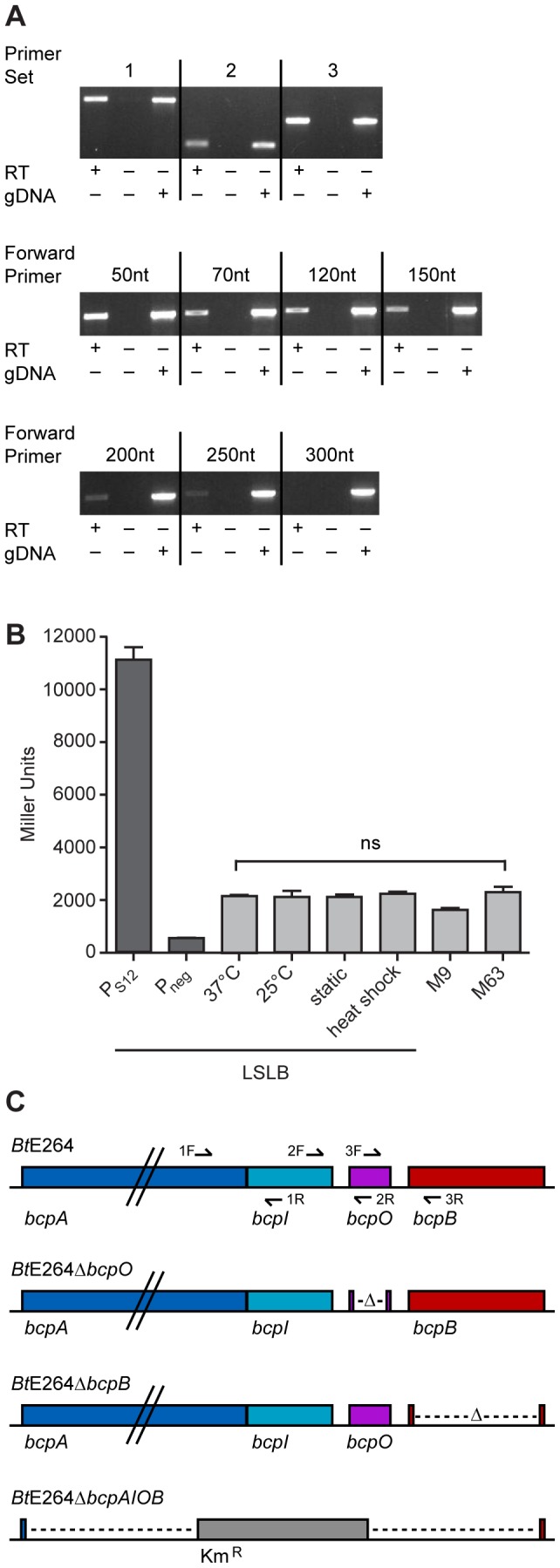
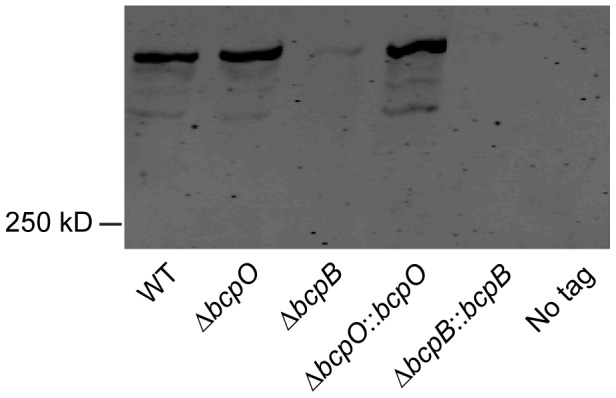
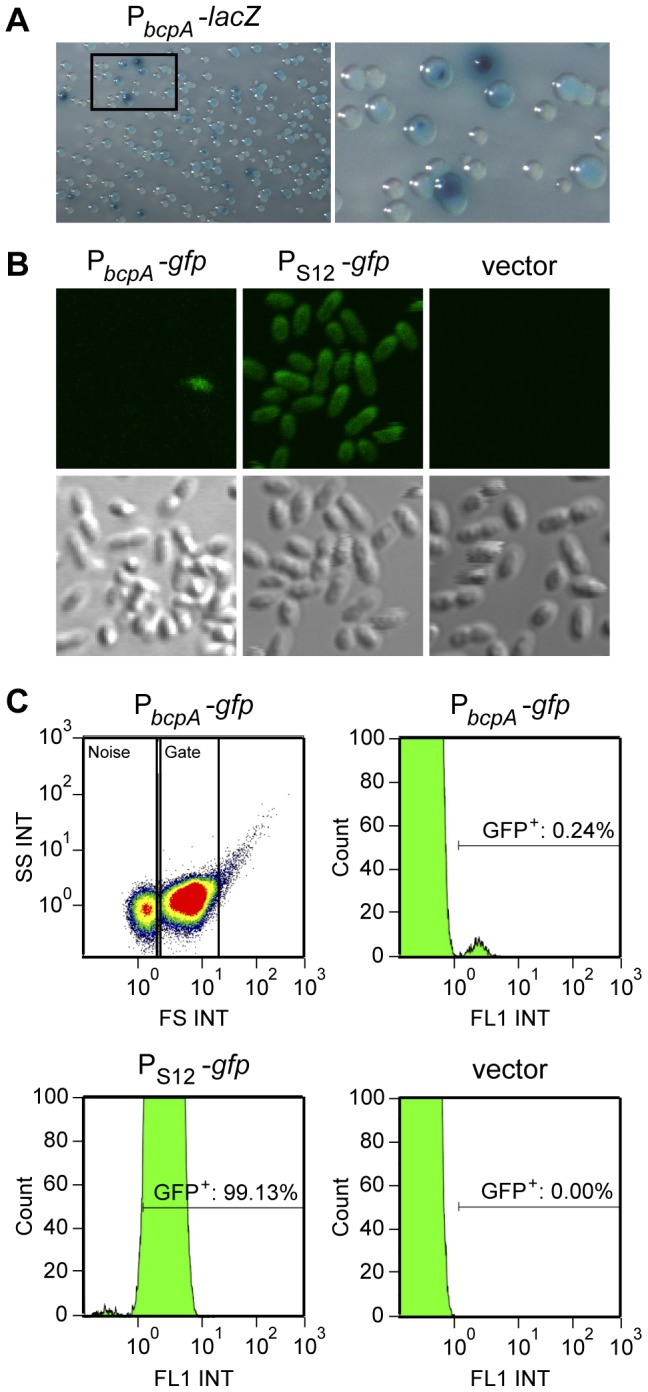
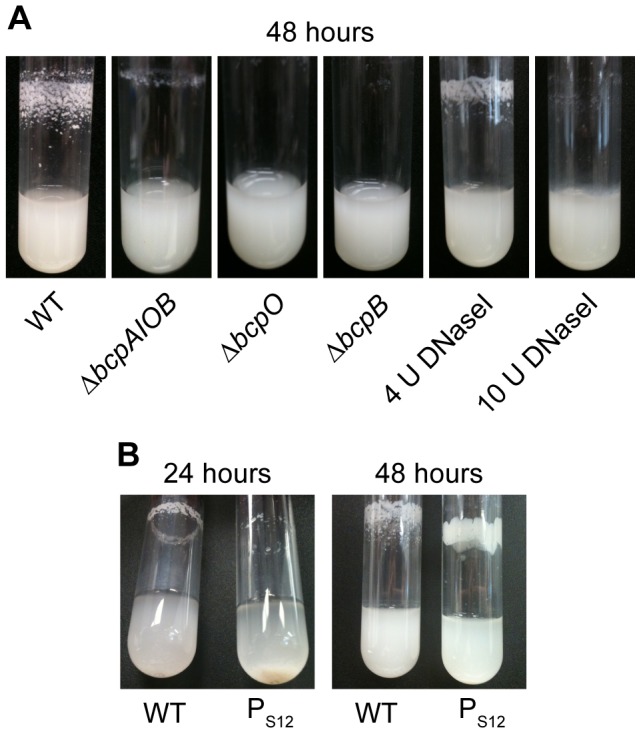
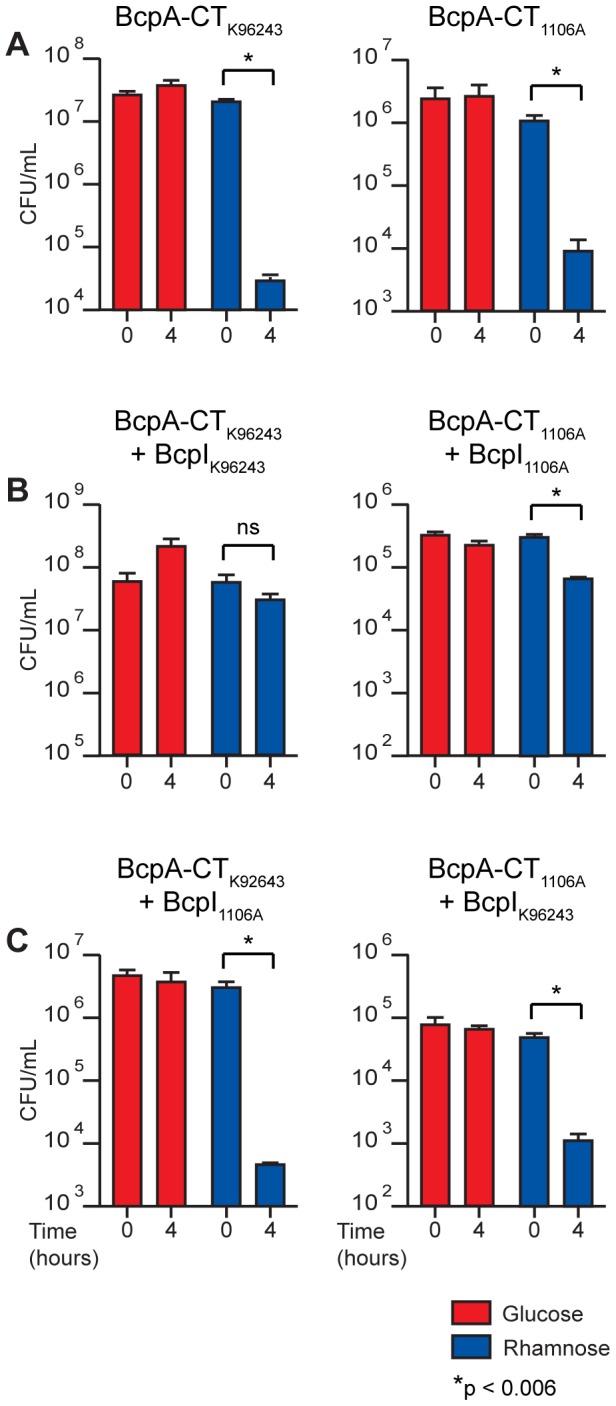
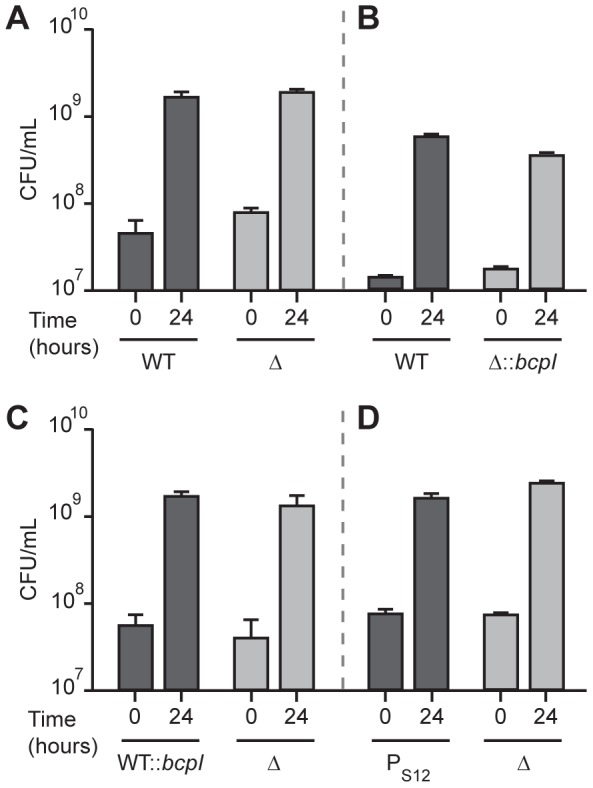
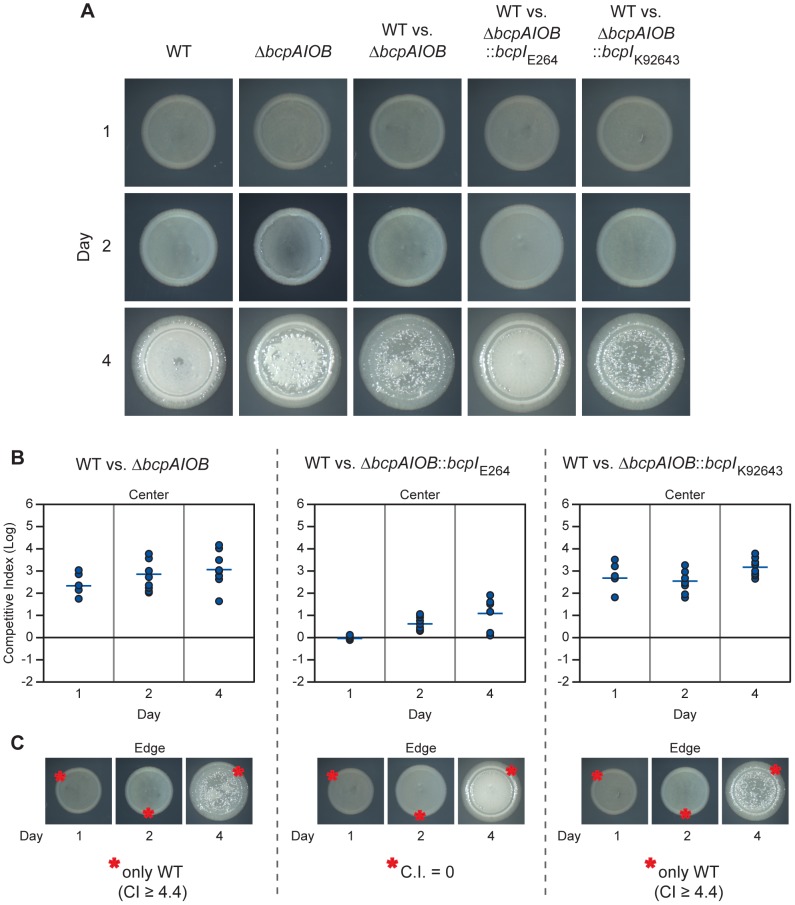
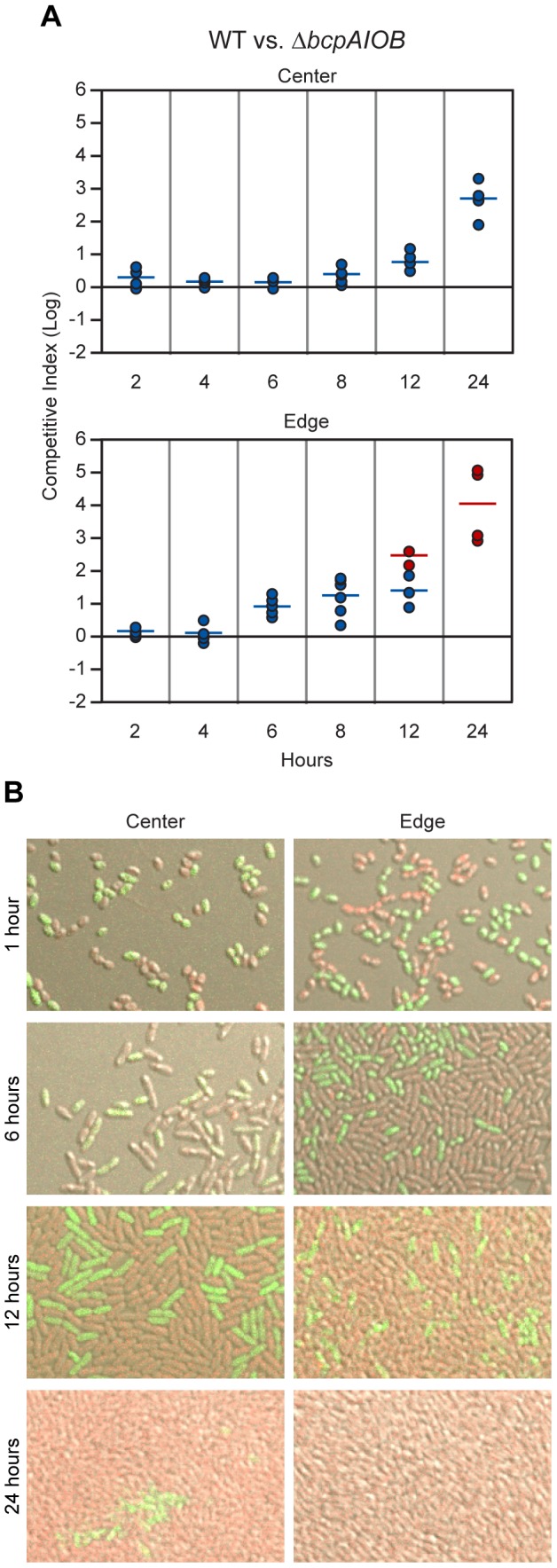
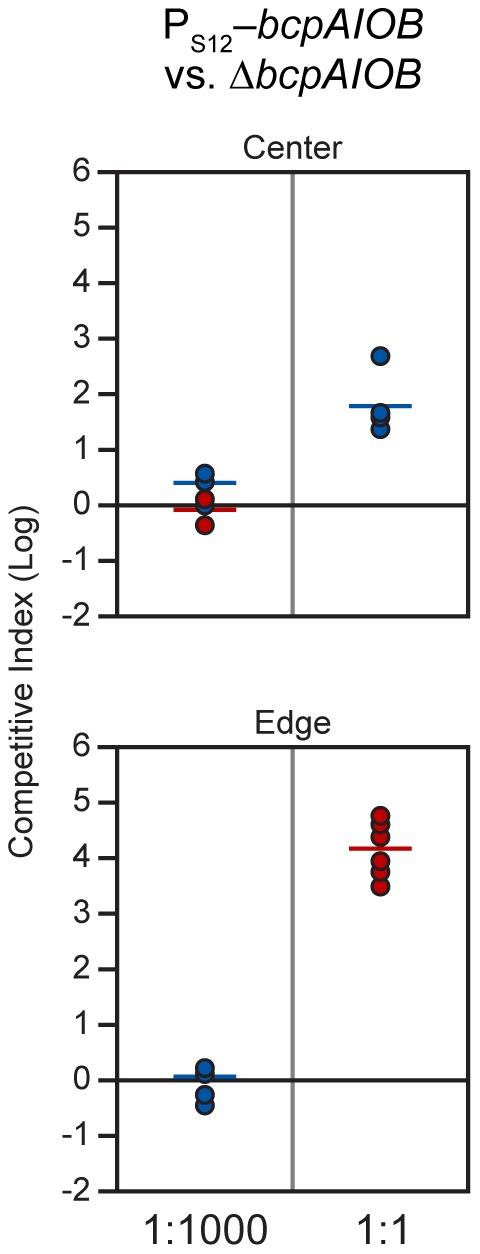
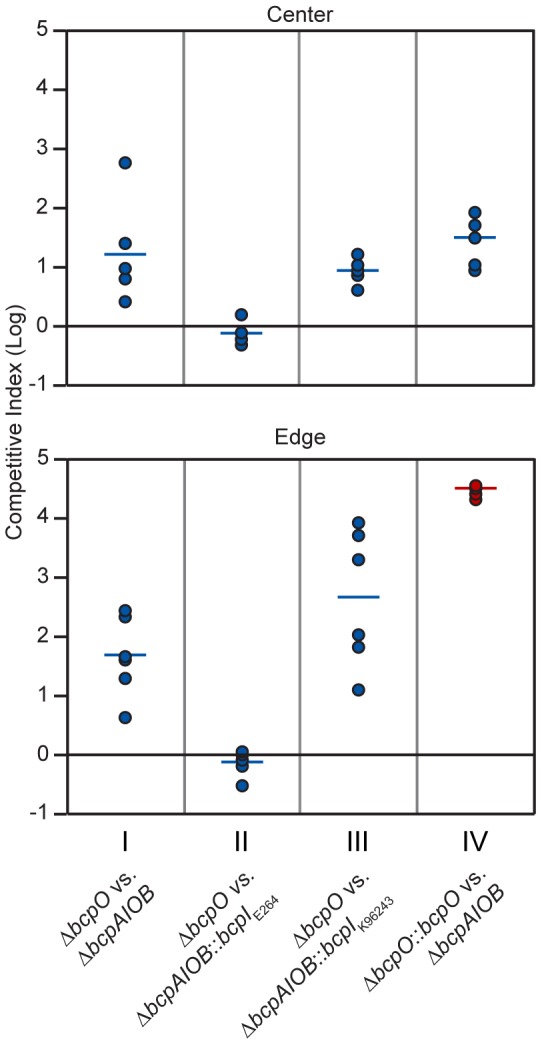
Microbes have evolved many strategies to adapt to changes in environmental conditions and population structures, including cooperation and competition. One apparently competitive mechanism is contact dependent growth inhibition (CDI). Identified in Escherichia coli, CDI is mediated by Two-Partner Secretion (TPS) pathway proteins, CdiA and CdiB. Upon cell contact, the toxic C-terminus of the TpsA family member CdiA, called the CdiA-CT, inhibits the growth of CDI(-) bacteria. CDI(+) bacteria are protected from autoinhibition by an immunity protein, CdiI. Bioinformatic analyses indicate that CDI systems are widespread amongst α, β, and γ proteobacteria and that the CdiA-CTs and CdiI proteins are highly variable. CdiI proteins protect against CDI in an allele-specific manner. Here we identify predicted CDI system-encoding loci in species of Burkholderia, Ralstonia and Cupriavidus, named bcpAIOB, that are distinguished from previously-described CDI systems by gene order and the presence of a small ORF, bcpO, located 5' to the gene encoding the TpsB family member. A requirement for bcpO in function of BcpA (the TpsA family member) was demonstrated, indicating that bcpAIOB define a novel class of TPS system. Using fluorescence microscopy and flow cytometry, we show that these genes are expressed in a probabilistic manner during culture of Burkholderia thailandensis in liquid medium. The bcpAIOB genes and extracellular DNA were required for autoaggregation and adherence to an abiotic surface, suggesting that CDI is required for biofilm formation, an activity not previously attributed to CDI. By contrast to what has been observed in E. coli, the B. thailandensis bcpAIOB genes only mediated interbacterial competition on a solid surface. Competition occurred in a defined spatiotemporal manner and was abrogated by allele-specific immunity. Our data indicate that the bcpAIOB genes encode distinct classes of CDI and TPS systems that appear to function in sociomicrobiological community development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Mechanisms and biological roles of contact-dependent growth inhibition systems.Cold Spring Harb Perspect Med. 2014 Feb 1;4(2):a010025. doi: 10.1101/cshperspect.a010025. Cold Spring Harb Perspect Med. 2014. PMID: 24492845 Free PMC article. Review.
-
Identification of functional toxin/immunity genes linked to contact-dependent growth inhibition (CDI) and rearrangement hotspot (Rhs) systems.PLoS Genet. 2011 Aug;7(8):e1002217. doi: 10.1371/journal.pgen.1002217. Epub 2011 Aug 4. PLoS Genet. 2011. PMID: 21829394 Free PMC article.
-
Kind discrimination and competitive exclusion mediated by contact-dependent growth inhibition systems shape biofilm community structure.PLoS Pathog. 2014 Apr 17;10(4):e1004076. doi: 10.1371/journal.ppat.1004076. eCollection 2014 Apr. PLoS Pathog. 2014. PMID: 24743836 Free PMC article.
-
A widespread family of polymorphic contact-dependent toxin delivery systems in bacteria.Nature. 2010 Nov 18;468(7322):439-42. doi: 10.1038/nature09490. Nature. 2010. PMID: 21085179 Free PMC article.
-
Contact-Dependent Growth Inhibition (CDI) and CdiB/CdiA Two-Partner Secretion Proteins.J Mol Biol. 2015 Nov 20;427(23):3754-65. doi: 10.1016/j.jmb.2015.09.010. Epub 2015 Sep 24. J Mol Biol. 2015. PMID: 26388411 Free PMC article. Review.
Cited by
-
Interbacterial signaling via Burkholderia contact-dependent growth inhibition system proteins.Proc Natl Acad Sci U S A. 2016 Jul 19;113(29):8296-301. doi: 10.1073/pnas.1606323113. Epub 2016 Jun 22. Proc Natl Acad Sci U S A. 2016. PMID: 27335458 Free PMC article.
-
Burkholderia BcpA mediates biofilm formation independently of interbacterial contact-dependent growth inhibition.Mol Microbiol. 2013 Sep;89(6):1213-25. doi: 10.1111/mmi.12339. Epub 2013 Aug 16. Mol Microbiol. 2013. PMID: 23879629 Free PMC article.
-
Bacterial contact-dependent growth inhibition.Trends Microbiol. 2013 May;21(5):230-7. doi: 10.1016/j.tim.2013.02.003. Epub 2013 Mar 7. Trends Microbiol. 2013. PMID: 23473845 Free PMC article. Review.
-
Mechanisms and biological roles of contact-dependent growth inhibition systems.Cold Spring Harb Perspect Med. 2014 Feb 1;4(2):a010025. doi: 10.1101/cshperspect.a010025. Cold Spring Harb Perspect Med. 2014. PMID: 24492845 Free PMC article. Review.
-
Proteus mirabilis Employs a Contact-Dependent Killing System against Competing Enterobacteriaceae.mSphere. 2021 Aug 25;6(4):e0032121. doi: 10.1128/mSphere.00321-21. Epub 2021 Jul 28. mSphere. 2021. PMID: 34319125 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
