

Phosphocreatine interacts with phospholipids, affects membrane properties and exerts membrane-protective effects
- PMID: 22912820
- PMCID: PMC3422282
- DOI: 10.1371/journal.pone.0043178
Phosphocreatine interacts with phospholipids, affects membrane properties and exerts membrane-protective effects
Abstract
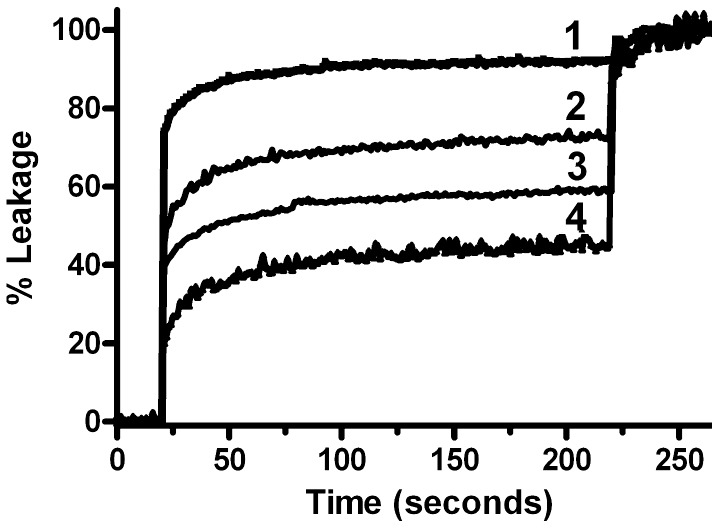
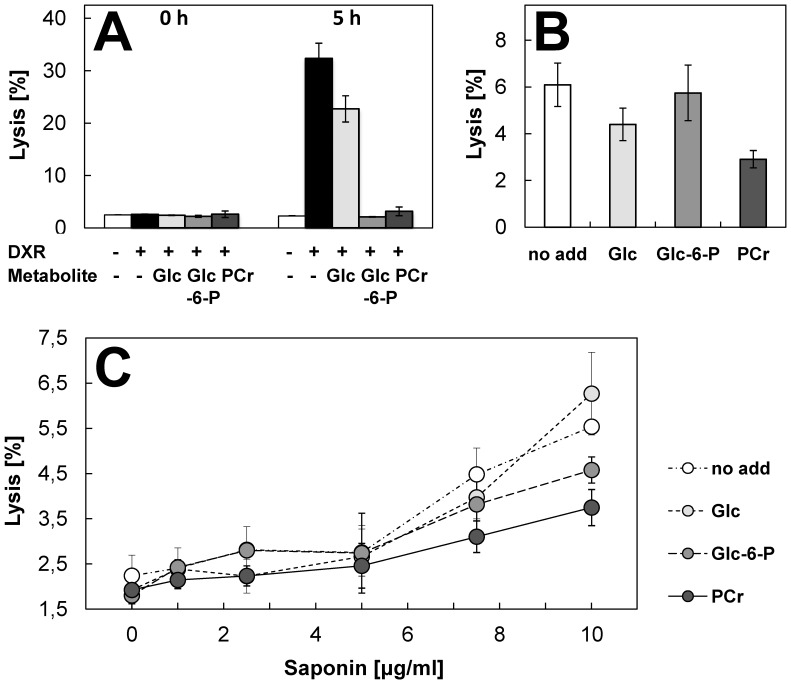
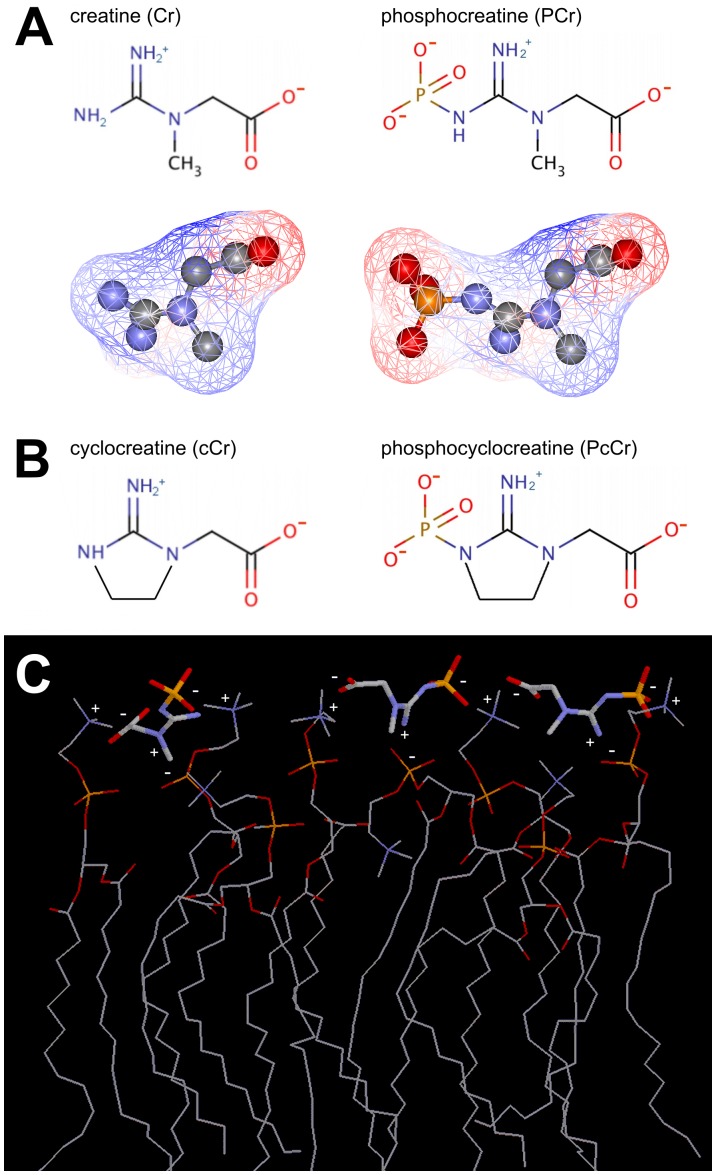
A broad spectrum of beneficial effects has been ascribed to creatine (Cr), phosphocreatine (PCr) and their cyclic analogues cyclo-(cCr) and phospho-cyclocreatine (PcCr). Cr is widely used as nutritional supplement in sports and increasingly also as adjuvant treatment for pathologies such as myopathies and a plethora of neurodegenerative diseases. Additionally, Cr and its cyclic analogues have been proposed for anti-cancer treatment. The mechanisms involved in these pleiotropic effects are still controversial and far from being understood. The reversible conversion of Cr and ATP into PCr and ADP by creatine kinase, generating highly diffusible PCr energy reserves, is certainly an important element. However, some protective effects of Cr and analogues cannot be satisfactorily explained solely by effects on the cellular energy state. Here we used mainly liposome model systems to provide evidence for interaction of PCr and PcCr with different zwitterionic phospholipids by applying four independent, complementary biochemical and biophysical assays: (i) chemical binding assay, (ii) surface plasmon resonance spectroscopy (SPR), (iii) solid-state (31)P-NMR, and (iv) differential scanning calorimetry (DSC). SPR revealed low affinity PCr/phospholipid interaction that additionally induced changes in liposome shape as indicated by NMR and SPR. Additionally, DSC revealed evidence for membrane packing effects by PCr, as seen by altered lipid phase transition. Finally, PCr efficiently protected against membrane permeabilization in two different model systems: liposome-permeabilization by the membrane-active peptide melittin, and erythrocyte hemolysis by the oxidative drug doxorubicin, hypoosmotic stress or the mild detergent saponin. These findings suggest a new molecular basis for non-energy related functions of PCr and its cyclic analogue. PCr/phospholipid interaction and alteration of membrane structure may not only protect cellular membranes against various insults, but could have more general implications for many physiological membrane-related functions that are relevant for health and disease.
Conflict of interest statement
Figures
Similar articles
-
Critical role of phosphagens in the energy cascade of cutaneous ischemia and protective action of phosphocreatine analogues in skin flap survival.Plast Reconstr Surg. 1998 May;101(6):1597-603. doi: 10.1097/00006534-199805000-00025. Plast Reconstr Surg. 1998. PMID: 9583491
-
Phosphocyclocreatine is the dominant form of cyclocreatine in control and creatine transporter deficiency patient fibroblasts.Pharmacol Res Perspect. 2019 Dec;7(6):e00525. doi: 10.1002/prp2.525. Pharmacol Res Perspect. 2019. PMID: 31859463 Free PMC article.
-
[Membranotropic effect of phosphocreatine and its structural analogs].Biokhimiia. 1991 Sep;56(9):1701-9. Biokhimiia. 1991. PMID: 1660733 Russian.
-
The creatine kinase system and pleiotropic effects of creatine.Amino Acids. 2011 May;40(5):1271-96. doi: 10.1007/s00726-011-0877-3. Epub 2011 Mar 30. Amino Acids. 2011. PMID: 21448658 Free PMC article. Review.
-
Systems bioenergetics of creatine kinase networks: physiological roles of creatine and phosphocreatine in regulation of cardiac cell function.Amino Acids. 2011 May;40(5):1333-48. doi: 10.1007/s00726-011-0854-x. Epub 2011 Mar 10. Amino Acids. 2011. PMID: 21390528 Review.
Cited by
-
Involvement of PI3K/Akt Signaling Pathway and Its Downstream Intracellular Targets in the Antidepressant-Like Effect of Creatine.Mol Neurobiol. 2016 Jul;53(5):2954-2968. doi: 10.1007/s12035-015-9192-4. Epub 2015 May 6. Mol Neurobiol. 2016. PMID: 25943184
-
Guanidinoacetic acid is efficacious in improving growth performance and muscle energy homeostasis in broiler chicks fed arginine-deficient or arginine-adequate diets.Poult Sci. 2019 Jul 1;98(7):2896-2905. doi: 10.3382/ps/pez036. Poult Sci. 2019. PMID: 30850832 Free PMC article.
-
Myocardial creatine levels do not influence response to acute oxidative stress in isolated perfused heart.PLoS One. 2014 Oct 1;9(10):e109021. doi: 10.1371/journal.pone.0109021. eCollection 2014. PLoS One. 2014. PMID: 25272153 Free PMC article.
-
High-Energy Phosphates and Ischemic Heart Disease: From Bench to Bedside.Front Cardiovasc Med. 2021 Jul 28;8:675608. doi: 10.3389/fcvm.2021.675608. eCollection 2021. Front Cardiovasc Med. 2021. PMID: 34395552 Free PMC article. Review.
-
Augmentation of Creatine in the Heart.Mini Rev Med Chem. 2016;16(1):19-28. doi: 10.2174/1389557515666150722102151. Mini Rev Med Chem. 2016. PMID: 26202199 Free PMC article. Review.
References
-
- Wyss M, Kaddurah-Daouk R (2000) Creatine and creatinine metabolism. Physiol Rev 80: 1107–1213. - PubMed
-
- Wallimann T, Tokarska-Schlattner M, Neumann D, Epand RM, Epand RF, et al... (2007) The phospho-creatine circuit: Molecular and cellular physiology of creatine kinases, sensitivity to free radicals and enhancement by creatine supplementation. In: Saks VA, editor. Molecular systems bioenergetics Energy for life. Weinheim: Wiley-VCH. 195–264.
-
- Wallimann T (2007) Introduction - creatine: cheap ergogenic supplement with great potential for health and disease. Subcell Biochem 46: 1–16. - PubMed
-
- Wyss M, Braissant O, Pischel I, Salomons GS, Schulze A, et al. (2007) Creatine and creatine kinase in health and disease - a bright future ahead? Subcell Biochem 46: 309–334. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
