

α-Conotoxin PeIA[S9H,V10A,E14N] potently and selectively blocks α6β2β3 versus α6β4 nicotinic acetylcholine receptors
- PMID: 22914547
- PMCID: PMC3477225
- DOI: 10.1124/mol.112.080853
α-Conotoxin PeIA[S9H,V10A,E14N] potently and selectively blocks α6β2β3 versus α6β4 nicotinic acetylcholine receptors
Abstract
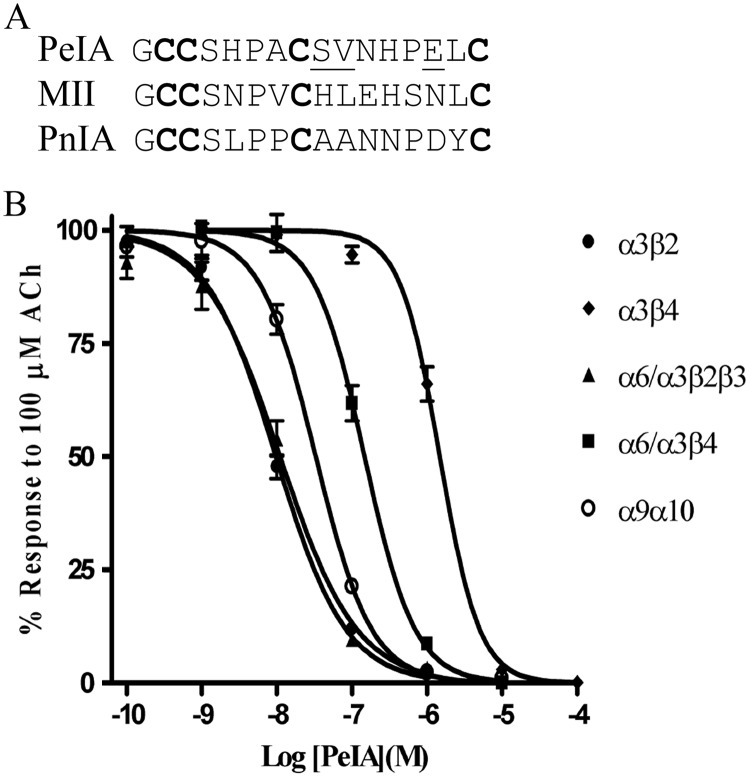
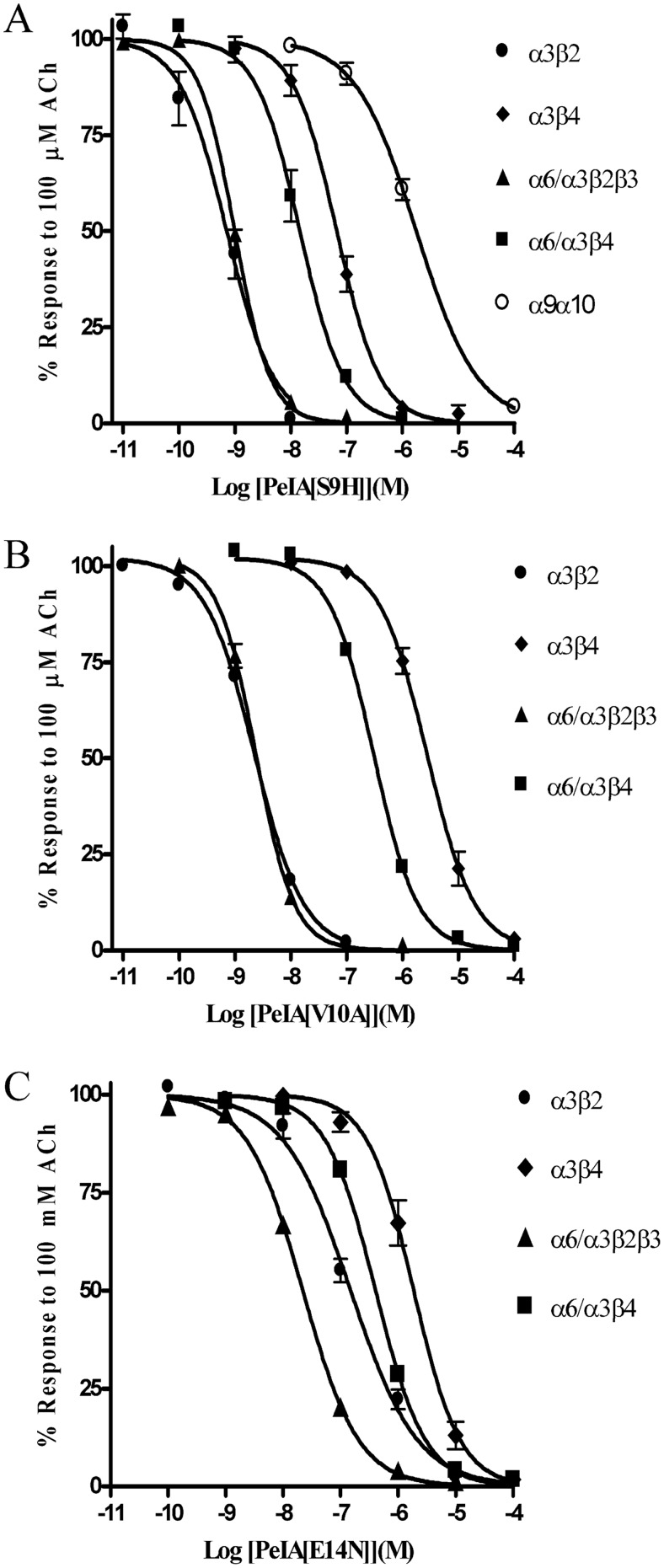
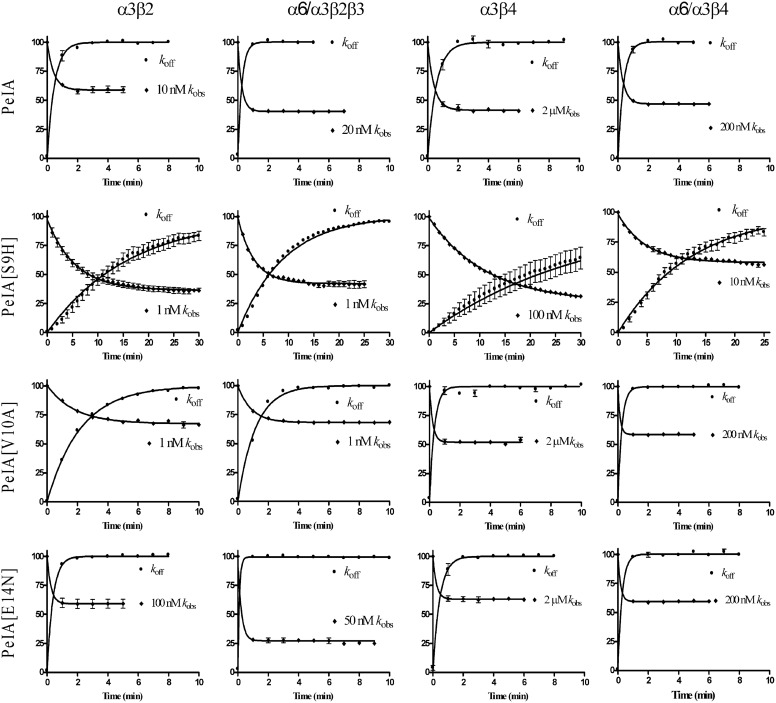
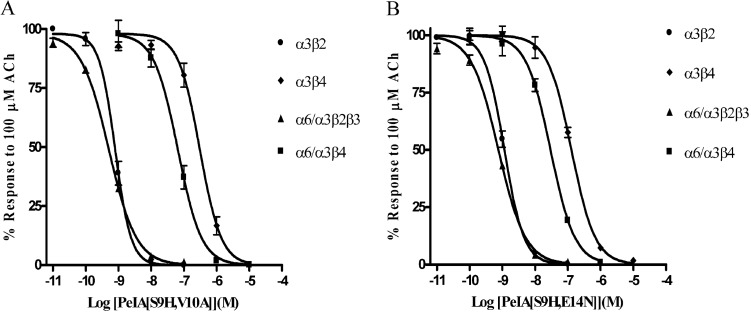
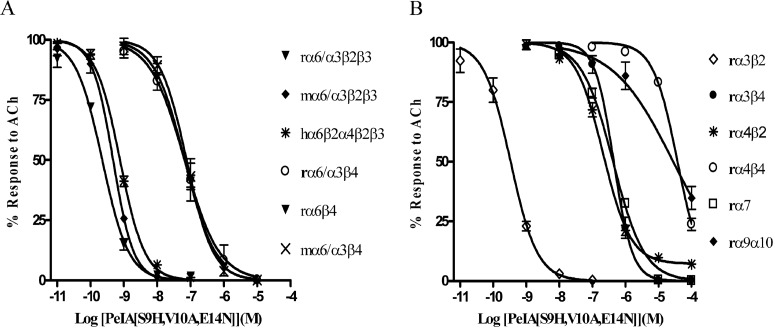
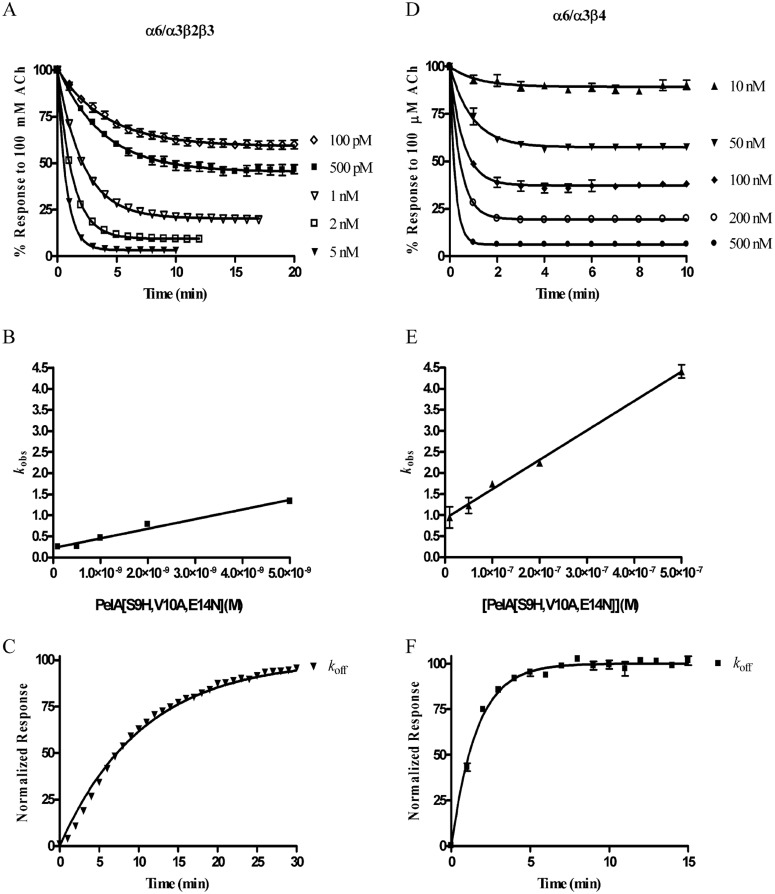
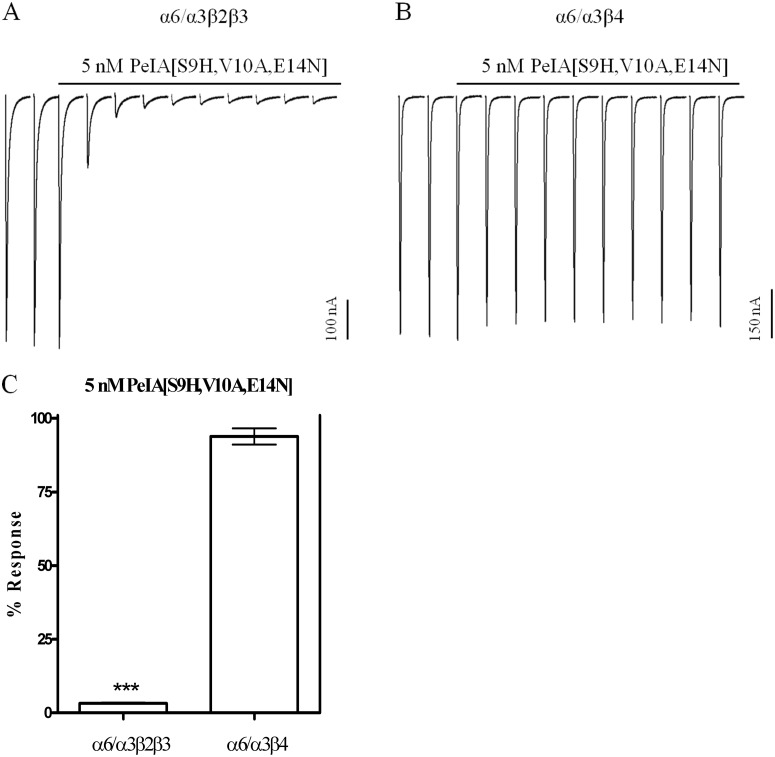
Nicotinic acetylcholine receptors (nAChRs) containing α6 and β2 subunits modulate dopamine release in the basal ganglia and are therapeutically relevant targets for treatment of neurological and psychiatric disorders including Parkinson's disease and nicotine dependence. However, the expression profile of β2 and β4 subunits overlap in a variety of tissues including locus ceruleus, retina, hippocampus, dorsal root ganglia, and adrenal chromaffin cells. Ligands that bind α6β2 nAChRs also potently bind the closely related α6β4 subtype. To distinguish between these two subtypes, we synthesized novel analogs of a recently described α-conotoxin, PeIA. PeIA is a peptide antagonist that blocks several nAChR subtypes, including α6/α3β2β3 and α6/α3β4 nAChRs, with low nanomolar potency. We systematically mutated PeIA and evaluated the resulting analogs for enhanced potency and/or selectivity for α6/α3β2β3 nAChRs expressed in Xenopus oocytes (α6/α3 is a subunit chimera that contains the N-terminal ligand-binding domain of the α6 subunit). On the basis of these results, second-generation analogs were then synthesized. The final analog, PeIA[S9H,V10A,E14N], potently blocked acetylcholine-gated currents mediated by α6/α3β2β3 and α6/α3β4 nAChRs with IC(50) values of 223 pM and 65 nM, respectively, yielding a >290-fold separation between the two subtypes. Kinetic studies of ligand binding to α6/α3β2β3 nAChRs yielded a k(off) of 0.096 ± 0.001 min(-1) and a k(on) of 0.23 ± 0.019 min(-1) M(-9). The synthesis of PeIA[S9H,V10A,E14N] demonstrates that ligands can be developed to discriminate between α6β2 and α6β4 nAChRs.
Figures
Similar articles
-
α-Conotoxins Identify the α3β4* Subtype as the Predominant Nicotinic Acetylcholine Receptor Expressed in Human Adrenal Chromaffin Cells.Mol Pharmacol. 2015 Nov;88(5):881-93. doi: 10.1124/mol.115.100982. Epub 2015 Sep 1. Mol Pharmacol. 2015. PMID: 26330550 Free PMC article.
-
α-Conotoxin VnIB from Conus ventricosus is a potent and selective antagonist of α6β4* nicotinic acetylcholine receptors.Neuropharmacology. 2019 Oct;157:107691. doi: 10.1016/j.neuropharm.2019.107691. Epub 2019 Jun 28. Neuropharmacology. 2019. PMID: 31255696 Free PMC article.
-
Positional scanning mutagenesis of α-conotoxin PeIA identifies critical residues that confer potency and selectivity for α6/α3β2β3 and α3β2 nicotinic acetylcholine receptors.J Biol Chem. 2013 Aug 30;288(35):25428-25439. doi: 10.1074/jbc.M113.482059. Epub 2013 Jul 11. J Biol Chem. 2013. PMID: 23846688 Free PMC article.
-
Mysterious alpha6-containing nAChRs: function, pharmacology, and pathophysiology.Acta Pharmacol Sin. 2009 Jun;30(6):740-51. doi: 10.1038/aps.2009.63. Acta Pharmacol Sin. 2009. PMID: 19498417 Free PMC article. Review.
-
The subtypes of nicotinic acetylcholine receptors on dopaminergic terminals of mouse striatum.Biochem Pharmacol. 2007 Oct 15;74(8):1235-46. doi: 10.1016/j.bcp.2007.07.032. Epub 2007 Jul 27. Biochem Pharmacol. 2007. PMID: 17825262 Free PMC article. Review.
Cited by
-
Qualitative Assay to Detect Dopamine Release by Ligand Action on Nicotinic Acetylcholine Receptors.Toxins (Basel). 2019 Nov 20;11(12):682. doi: 10.3390/toxins11120682. Toxins (Basel). 2019. PMID: 31757080 Free PMC article.
-
Molecular determinants of α-conotoxin potency for inhibition of human and rat α6β4 nicotinic acetylcholine receptors.J Biol Chem. 2018 Nov 16;293(46):17838-17852. doi: 10.1074/jbc.RA118.005649. Epub 2018 Sep 24. J Biol Chem. 2018. PMID: 30249616 Free PMC article.
-
Expression of α3β2β4 nicotinic acetylcholine receptors by rat adrenal chromaffin cells determined using novel conopeptide antagonists.J Neurochem. 2020 Jul;154(2):158-176. doi: 10.1111/jnc.14966. Epub 2020 Feb 11. J Neurochem. 2020. PMID: 31967330 Free PMC article.
-
Conotoxins targeting nicotinic acetylcholine receptors: an overview.Mar Drugs. 2014 May 22;12(5):2970-3004. doi: 10.3390/md12052970. Mar Drugs. 2014. PMID: 24857959 Free PMC article. Review.
-
Prediction of Specificity of α-Conotoxins to Subtypes of Human Nicotinic Acetylcholine Receptors with Semi-supervised Machine Learning.ACS Chem Neurosci. 2025 Jun 18;16(12):2196-2207. doi: 10.1021/acschemneuro.4c00760. Epub 2025 May 29. ACS Chem Neurosci. 2025. PMID: 40441695 Free PMC article.
References
-
- Azam L, McIntosh JM. (2006) Characterization of nicotinic acetylcholine receptors that modulate nicotine-evoked [3H]norepinephrine release from mouse hippocampal synaptosomes. Mol Pharmacol 70:967–976 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
