

Glycan-dependent immunogenicity of recombinant soluble trimeric hemagglutinin
- PMID: 22915811
- PMCID: PMC3486279
- DOI: 10.1128/JVI.01084-12
Glycan-dependent immunogenicity of recombinant soluble trimeric hemagglutinin
Abstract
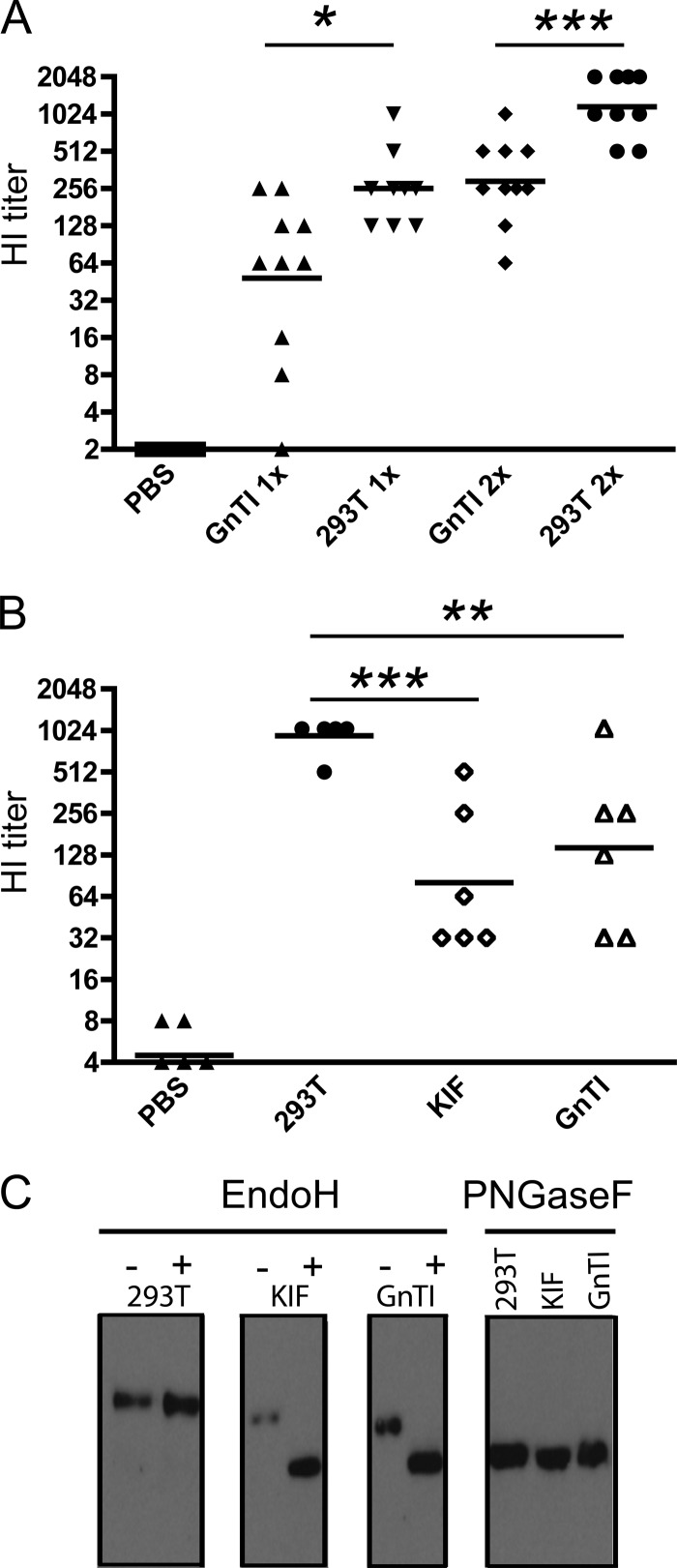
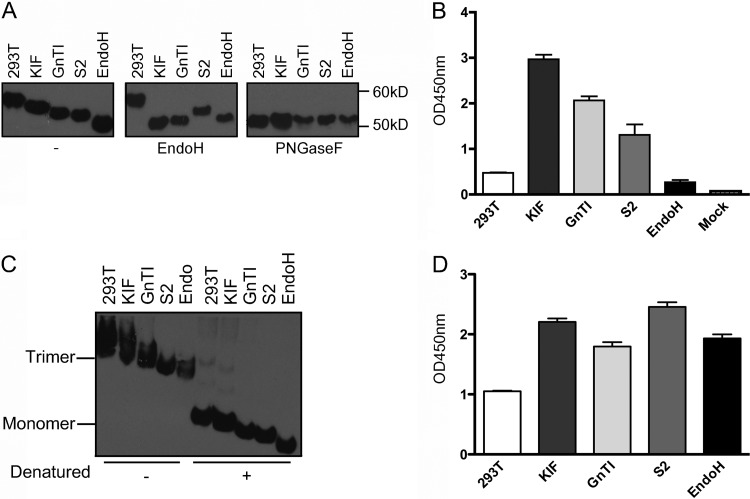
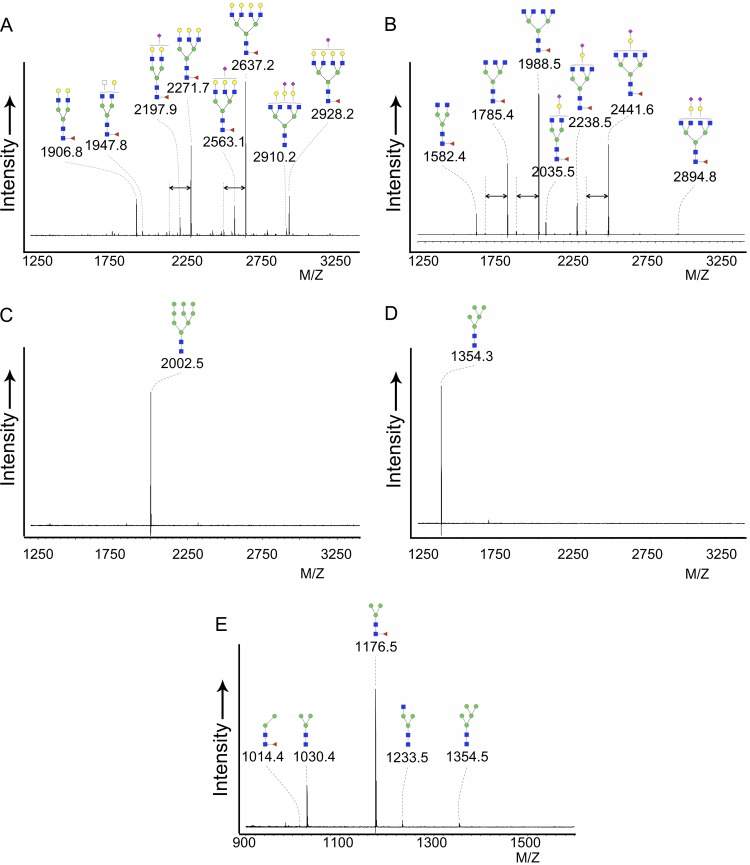
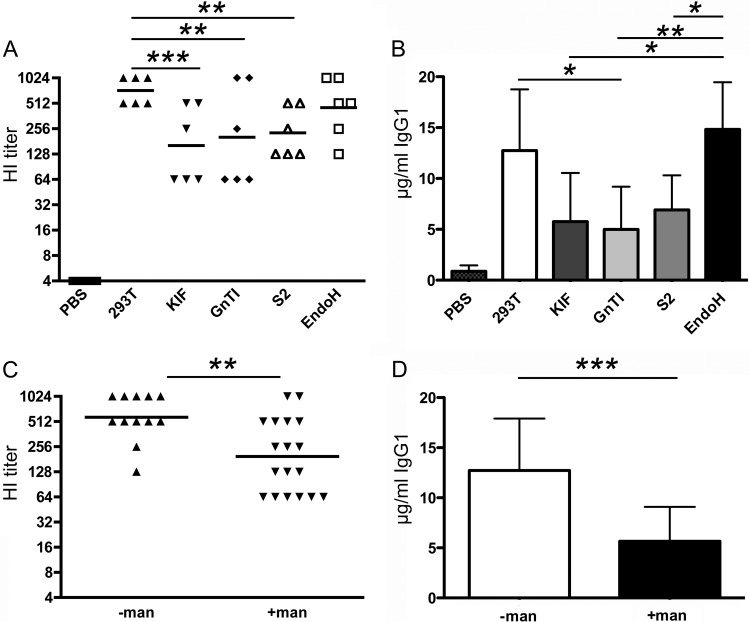
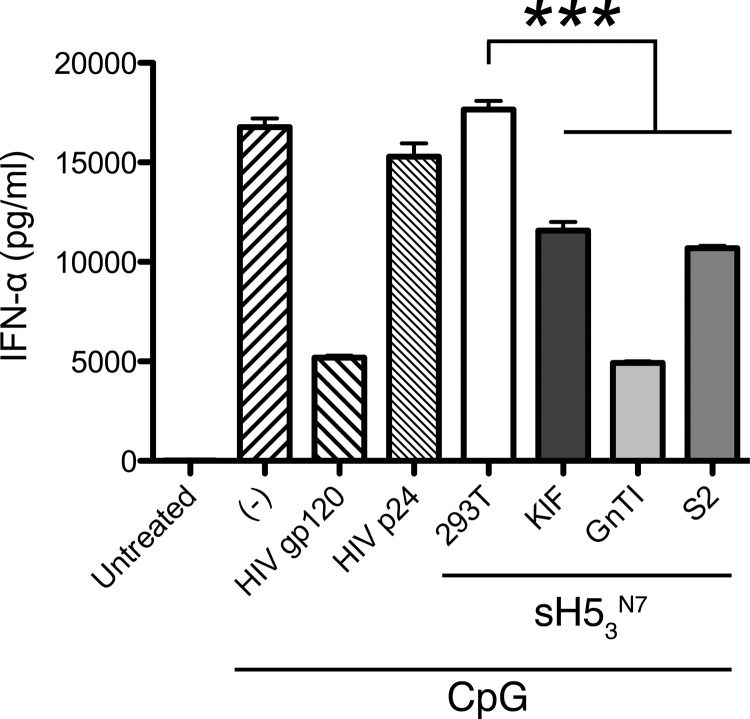
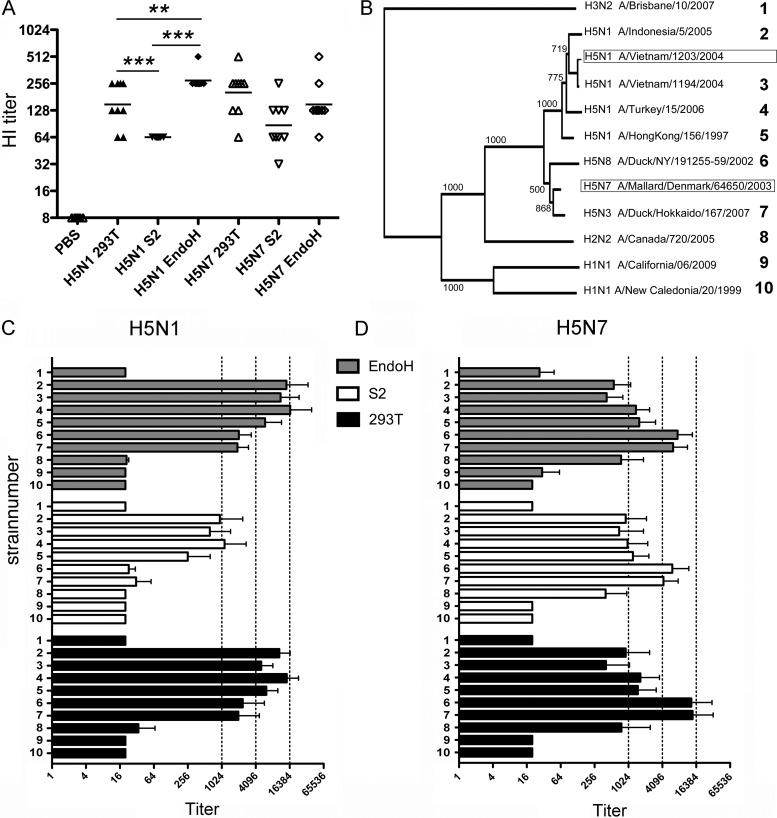
Recombinant soluble trimeric influenza A virus (IAV) hemagglutinin (sHA(3)) has proven an effective vaccine antigen against IAV. Here, we investigate to what extent the glycosylation status of the sHA(3) glycoprotein affects its immunogenicity. Different glycosylation forms of subtype H5 trimeric HA protein (sH5(3)) were produced by expression in insect cells and different mammalian cells in the absence and presence of inhibitors of N-glycan-modifying enzymes or by enzymatic removal of the oligosaccharides. The following sH5(3) preparations were evaluated: (i) HA proteins carrying complex glycans produced in HEK293T cells; (ii) HA proteins carrying Man(9)GlcNAc(2) moieties, expressed in HEK293T cells treated with kifunensine; (iii) HA proteins containing Man(5)GlcNAc(2) moieties derived from HEK293S GnTI(-) cells; (iv) insect cell-produced HA proteins carrying paucimannosidic N-glycans; and (v) HEK293S GnTI(-) cell-produced HA proteins treated with endoglycosidase H, thus carrying side chains composed of only a single N-acetylglucosamine each. The different HA glycosylation states were confirmed by comparative electrophoretic analysis and by mass spectrometric analysis of released glycans. The immunogenicity of the HA preparations was studied in chickens and mice. The results demonstrate that HA proteins carrying terminal mannose moieties induce significantly lower hemagglutination inhibition antibody titers than HA proteins carrying complex glycans or single N-acetylglucosamine side chains. However, the glycosylation state of the HA proteins did not affect the breadth of the antibody response as measured by an HA1 antigen microarray. We conclude that the glycosylation state of recombinant antigens is a factor of significant importance when developing glycoprotein-based vaccines, such as recombinant HA proteins.
Figures
Similar articles
-
Different immunity elicited by recombinant H5N1 hemagglutinin proteins containing pauci-mannose, high-mannose, or complex type N-glycans.PLoS One. 2013 Jun 14;8(6):e66719. doi: 10.1371/journal.pone.0066719. Print 2013. PLoS One. 2013. PMID: 23799128 Free PMC article.
-
The influenza A virus hemagglutinin glycosylation state affects receptor-binding specificity.Virology. 2010 Jul 20;403(1):17-25. doi: 10.1016/j.virol.2010.03.047. Epub 2010 May 2. Virology. 2010. PMID: 20441997
-
Influenza Virus Hemagglutinin Glycoproteins with Different N-Glycan Patterns Activate Dendritic Cells In Vitro.J Virol. 2016 Jun 10;90(13):6085-6096. doi: 10.1128/JVI.00452-16. Print 2016 Jul 1. J Virol. 2016. PMID: 27099319 Free PMC article.
-
Influenza Virus Hemagglutinins H2, H5, H6, and H11 Are Not Targets of Pulmonary Surfactant Protein D: N-Glycan Subtypes in Host-Pathogen Interactions.J Virol. 2020 Feb 14;94(5):e01951-19. doi: 10.1128/JVI.01951-19. Print 2020 Feb 14. J Virol. 2020. PMID: 31826991 Free PMC article.
-
Unmasking Stem-Specific Neutralizing Epitopes by Abolishing N-Linked Glycosylation Sites of Influenza Virus Hemagglutinin Proteins for Vaccine Design.J Virol. 2016 Sep 12;90(19):8496-508. doi: 10.1128/JVI.00880-16. Print 2016 Oct 1. J Virol. 2016. PMID: 27440889 Free PMC article.
Cited by
-
Impact of Protein Glycosylation on the Design of Viral Vaccines.Adv Biochem Eng Biotechnol. 2021;175:319-354. doi: 10.1007/10_2020_132. Adv Biochem Eng Biotechnol. 2021. PMID: 32935143 Review.
-
Influenza A Virus Agnostic Receptor Tropism Revealed Using a Novel Biological System with Terminal Sialic Acid Knockout Cells.J Virol. 2022 Aug 10;96(15):e0041622. doi: 10.1128/jvi.00416-22. Epub 2022 Jul 18. J Virol. 2022. PMID: 35862707 Free PMC article.
-
Prime-Boost Vaccination With a Novel Hemagglutinin Protein Produced in Bacteria Induces Neutralizing Antibody Responses Against H5-Subtype Influenza Viruses in Commercial Chickens.Front Immunol. 2019 Sep 4;10:2006. doi: 10.3389/fimmu.2019.02006. eCollection 2019. Front Immunol. 2019. PMID: 31552018 Free PMC article.
-
The Coming Age of Insect Cells for Manufacturing and Development of Protein Therapeutics.Ind Eng Chem Res. 2018 Aug 8;57(31):10061-10070. doi: 10.1021/acs.iecr.8b00985. Epub 2018 Jul 9. Ind Eng Chem Res. 2018. PMID: 30886455 Free PMC article.
-
Different immunity elicited by recombinant H5N1 hemagglutinin proteins containing pauci-mannose, high-mannose, or complex type N-glycans.PLoS One. 2013 Jun 14;8(6):e66719. doi: 10.1371/journal.pone.0066719. Print 2013. PLoS One. 2013. PMID: 23799128 Free PMC article.
References
-
- Cornelissen LA, et al. 2010. A single immunization with soluble recombinant trimeric hemagglutinin protects chickens against highly pathogenic avian influenza virus H5N1. PLoS One 5:e10645 doi:10.1371/journal.pone.0010645 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
