

Essential role of brain-derived neurotrophic factor in the regulation of serotonin transmission in the basolateral amygdala
- PMID: 22917617
- PMCID: PMC3475413
- DOI: 10.1016/j.neuroscience.2012.08.025
Essential role of brain-derived neurotrophic factor in the regulation of serotonin transmission in the basolateral amygdala
Abstract
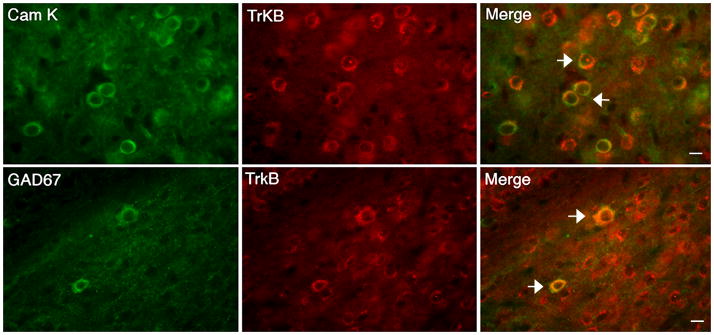
Human and animal model studies have linked brain-derived neurotrophic factor (BDNF) with the etiology of anxiety disorders. This pleiotropic neurotrophin and its receptor, TrkB, promote neuronal survival, differentiation and synaptic plasticity. Here we interrogated the role of BDNF in serotonergic neurotransmission in the basolateral amygdala (BLA), a limbic brain region associated with the neurobiology of anxiety. We found that both GABAergic and pyramidal projection neurons in the wild-type BLA contained TrkB receptors. Examination of BDNF(2L/2LCk-Cre) mutant mice with brain-selective depletion of BDNF revealed mild decreases in serotonin content in the BLA. Notably, whole cell recordings in BLA pyramidal cells uncovered significant alterations in 5-HT(2)-mediated regulation of GABAergic and glutamatergic transmission in BDNF(2L/2LCk-Cre) mutant mice that result in a hyperexcitable circuit. These changes were associated with decreased expression of 5-HT(2) receptors. Collectively, the results indicate a required role of BDNF in serotonin transmission in the BLA. Furthermore, they suggest a mechanism underlying the reported increase in anxiety-like behavior elicited by perturbed BDNF signaling.
Copyright © 2012 IBRO. Published by Elsevier Ltd. All rights reserved.
Figures






Similar articles
-
Behavioral and serotonergic consequences of decreasing or increasing hippocampus brain-derived neurotrophic factor protein levels in mice.Neuropharmacology. 2008 Nov;55(6):1006-14. doi: 10.1016/j.neuropharm.2008.08.001. Epub 2008 Aug 12. Neuropharmacology. 2008. PMID: 18761360 Review.
-
Impact of Chronic BDNF Depletion on GABAergic Synaptic Transmission in the Lateral Amygdala.Int J Mol Sci. 2019 Sep 3;20(17):4310. doi: 10.3390/ijms20174310. Int J Mol Sci. 2019. PMID: 31484392 Free PMC article.
-
Impaired GABAergic inhibition in the visual cortex of brain-derived neurotrophic factor heterozygous knockout mice.J Physiol. 2008 Apr 1;586(7):1885-901. doi: 10.1113/jphysiol.2007.148627. Epub 2008 Jan 31. J Physiol. 2008. PMID: 18238806 Free PMC article.
-
Evidence for low GluR2 AMPA receptor subunit expression at synapses in the rat basolateral amygdala.J Neurochem. 2005 Sep;94(6):1728-38. doi: 10.1111/j.1471-4159.2005.03334.x. Epub 2005 Jul 25. J Neurochem. 2005. PMID: 16045445 Free PMC article.
-
Interaction between BDNF and serotonin: role in mood disorders.Neuropsychopharmacology. 2008 Jan;33(1):73-83. doi: 10.1038/sj.npp.1301571. Epub 2007 Sep 19. Neuropsychopharmacology. 2008. PMID: 17882234 Review.
Cited by
-
Aberrant Amygdala-Dependent Cued Fear Memory in Na+/Ca2+ Exchanger 1 Heterozygous Mice.Mol Neurobiol. 2019 Jun;56(6):4381-4394. doi: 10.1007/s12035-018-1384-2. Epub 2018 Oct 15. Mol Neurobiol. 2019. PMID: 30324228
-
Network Pharmacology Combined with Animal Models to Investigate the Mechanism of ChangPu YuJin Tang in the Treatment of Tourette Syndrome.Comb Chem High Throughput Screen. 2025;28(1):166-184. doi: 10.2174/0113862073295447240430113053. Comb Chem High Throughput Screen. 2025. PMID: 38706359 Free PMC article.
-
Beneficial effect of fluoxetine treatment aganist psychological stress is mediated by increasing BDNF expression in selected brain areas.Oncotarget. 2017 May 15;8(41):69527-69537. doi: 10.18632/oncotarget.17891. eCollection 2017 Sep 19. Oncotarget. 2017. PMID: 29050222 Free PMC article.
-
Increased brain-derived neurotrophic factor (BDNF) protein concentrations in mice lacking brain serotonin.Eur Arch Psychiatry Clin Neurosci. 2016 Apr;266(3):281-4. doi: 10.1007/s00406-015-0611-3. Epub 2015 Jun 23. Eur Arch Psychiatry Clin Neurosci. 2016. PMID: 26100147
-
Reduced levels of brain-derived neurotrophic factor contribute to synaptic imbalance during the critical period of respiratory development in rats.Eur J Neurosci. 2014 Jul;40(1):2183-95. doi: 10.1111/ejn.12568. Epub 2014 Mar 26. Eur J Neurosci. 2014. PMID: 24666389 Free PMC article.
References
-
- Aghajanian GK, Marek GJ. Serotonin induces excitatory postsynaptic potentials in apical dendrites of neocortical pyramidal cells. Neuropharmacology. 1997;36:589–599. - PubMed
-
- Bath KG, Chuang J, Spencer-Segal JL, Amso D, Altemus M, McEwen BS, Lee FS. Variant Brain Derived Neurotrophic Factor (Valine 66Methionine) Polymorphism Contributes to Developmental and Estrous Stage-Specific Expression of Anxiety-Like Behavior in Female Mice. Biological Psychiatry. 2012 May; (epub ahead of print) - PMC - PubMed
-
- Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, Graham D, Tsankova NM, Bolanos CA, Rios M, Monteggia LM, Self DW, Nestler EJ. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science. 2006;311(5762):864–8. - PubMed
-
- Bremner JD, Vermetten E, Schmahl C, Vaccarino V, Vythilingam M, Afzal N, Grillon C, Charney DS. Positron emission tomographic imaging of neural correlates of a fear acquisition and extinction paradigm in women with childhood sexual-abuse-related post-traumatic stress disorder. Psychol Med. 2005;35:791–806. - PMC - PubMed
-
- Bryant RA, Felmingham K, Kemp A, Das P, Hughes G, Peduto A, Williams L. Amygdala and ventral anterior cingulate activation predicts treatment response to cognitive behaviour therapy for post-traumatic stress disorder. Psychol Med. 2008;38:555–561. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
