

The wonders of flap endonucleases: structure, function, mechanism and regulation
- PMID: 22918592
- PMCID: PMC3728657
- DOI: 10.1007/978-94-007-4572-8_16
The wonders of flap endonucleases: structure, function, mechanism and regulation
Abstract
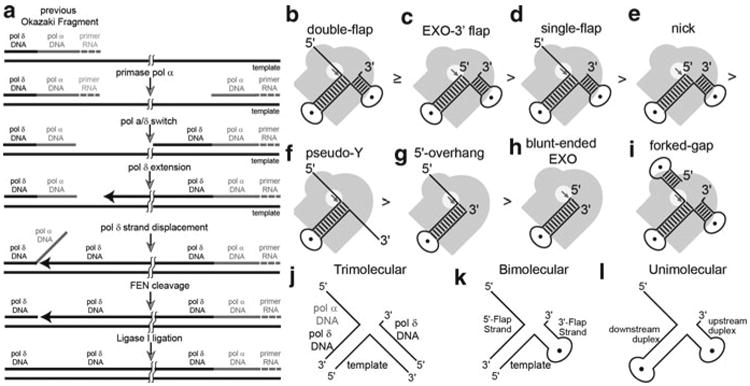
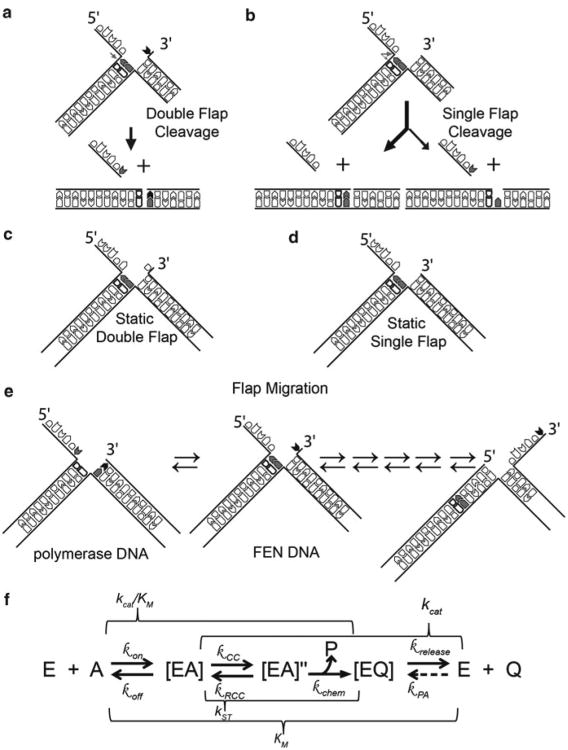
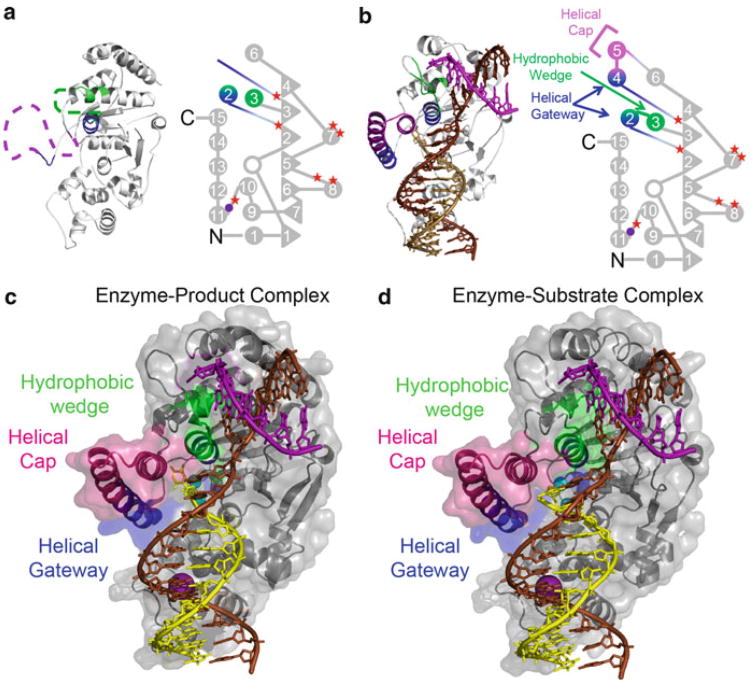
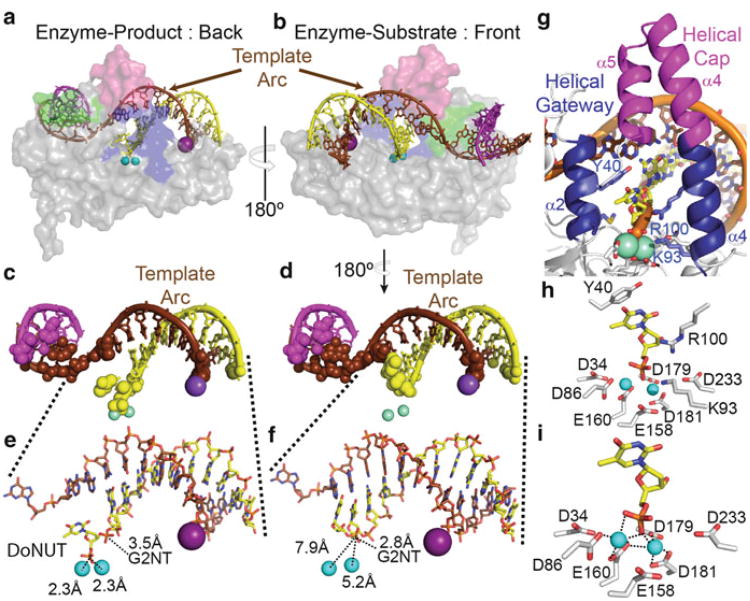
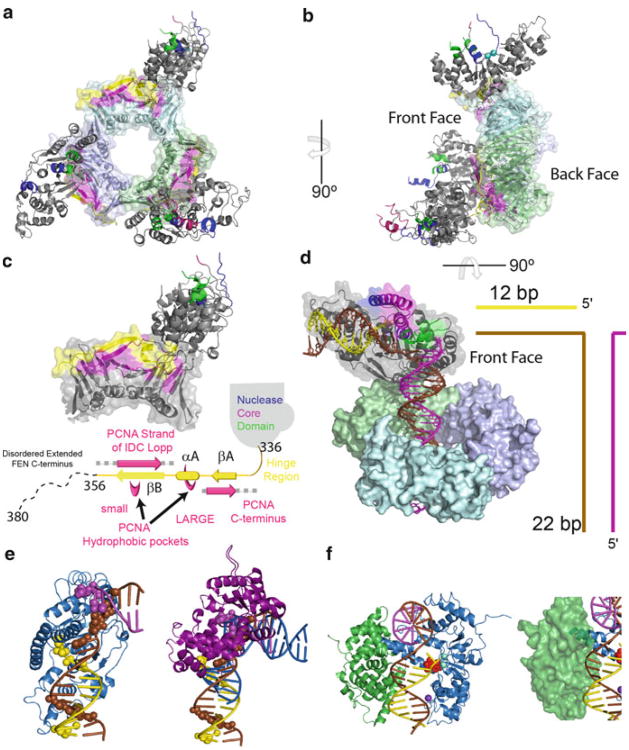
Processing of Okazaki fragments to complete lagging strand DNA synthesis requires coordination among several proteins. RNA primers and DNA synthesised by DNA polymerase α are displaced by DNA polymerase δ to create bifurcated nucleic acid structures known as 5'-flaps. These 5'-flaps are removed by Flap Endonuclease 1 (FEN), a structure-specific nuclease whose divalent metal ion-dependent phosphodiesterase activity cleaves 5'-flaps with exquisite specificity. FENs are paradigms for the 5' nuclease superfamily, whose members perform a wide variety of roles in nucleic acid metabolism using a similar nuclease core domain that displays common biochemical properties and structural features. A detailed review of FEN structure is undertaken to show how DNA substrate recognition occurs and how FEN achieves cleavage at a single phosphate diester. A proposed double nucleotide unpairing trap (DoNUT) is discussed with regards to FEN and has relevance to the wider 5' nuclease superfamily. The homotrimeric proliferating cell nuclear antigen protein (PCNA) coordinates the actions of DNA polymerase, FEN and DNA ligase by facilitating the hand-off intermediates between each protein during Okazaki fragment maturation to maximise through-put and minimise consequences of intermediates being released into the wider cellular environment. FEN has numerous partner proteins that modulate and control its action during DNA replication and is also controlled by several post-translational modification events, all acting in concert to maintain precise and appropriate cleavage of Okazaki fragment intermediates during DNA replication.
Figures
References
-
- Beattie TR, Bell SD. The role of the DNA sliding clamp in Okazaki fragment maturation in archaea and eukaryotes. Biochem Soc Trans. 2011;39:70–76. - PubMed
-
- Berg OG, von Hippel PH. Diffusion-controlled macromolecular interactions. Annu Rev Biophys Biophys Chem. 1985;14:131–160. - PubMed
-
- Bornarth CJ, Ranalli TA, Henricksen LA, Wahl AF, Bambara RA. Effect of flap modifications on human FEN1 cleavage. Biochemistry. 1999;38:13347–13354. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
