

Cdc42p regulation of the yeast formin Bni1p mediated by the effector Gic2p
- PMID: 22918946
- PMCID: PMC3459858
- DOI: 10.1091/mbc.E12-05-0400
Cdc42p regulation of the yeast formin Bni1p mediated by the effector Gic2p
Abstract
Actin filaments are dynamically reorganized to accommodate ever-changing cellular needs for intracellular transport, morphogenesis, and migration. Formins, a major family of actin nucleators, are believed to function as direct effectors of Rho GTPases, such as the polarity regulator Cdc42p. However, the presence of extensive redundancy has made it difficult to assess the in vivo significance of the low-affinity Rho GTPase-formin interaction and specifically whether Cdc42p polarizes the actin cytoskeleton via direct formin binding. Here we exploit a synthetically rewired budding yeast strain to eliminate the redundancy, making regulation of the formin Bni1p by Cdc42p essential for viability. Surprisingly, we find that direct Cdc42p-Bni1p interaction is dispensable for Bni1p regulation. Alternative paths linking Cdc42p and Bni1p via "polarisome" components Spa2p and Bud6p are also collectively dispensable. We identify a novel regulatory input to Bni1p acting through the Cdc42p effector, Gic2p. This pathway is sufficient to localize Bni1p to the sites of Cdc42p action and promotes a polarized actin organization in both rewired and wild-type contexts. We suggest that an indirect mechanism linking Rho GTPases and formins via Rho effectors may provide finer spatiotemporal control for the formin-nucleated actin cytoskeleton.
Figures
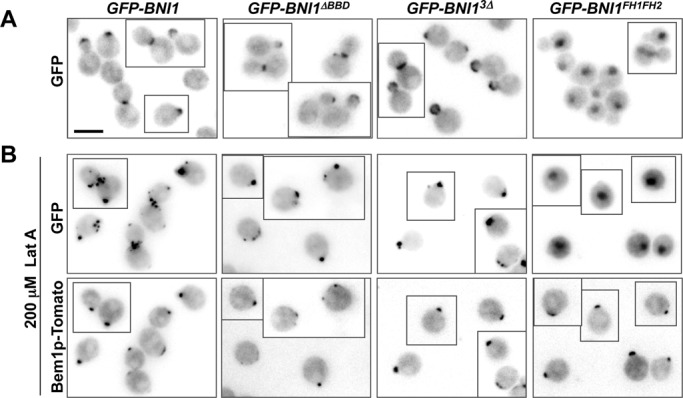
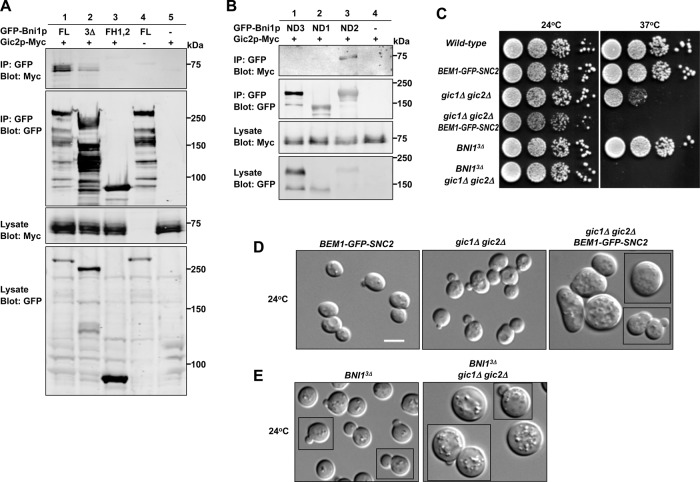
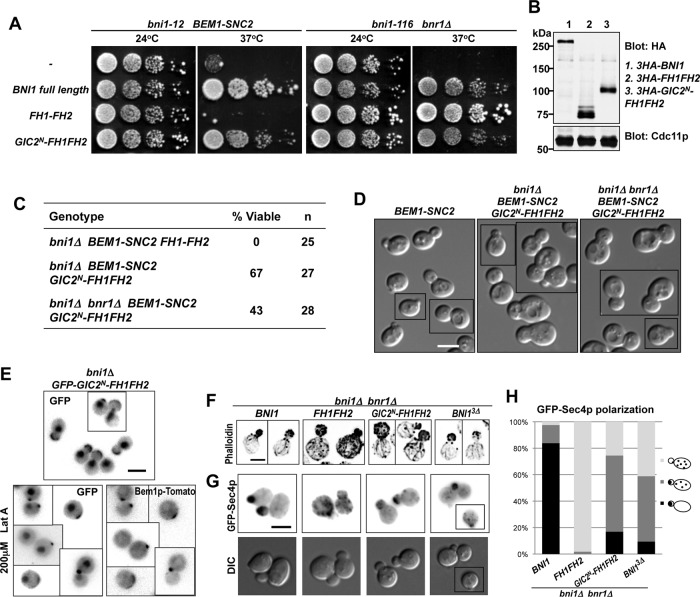
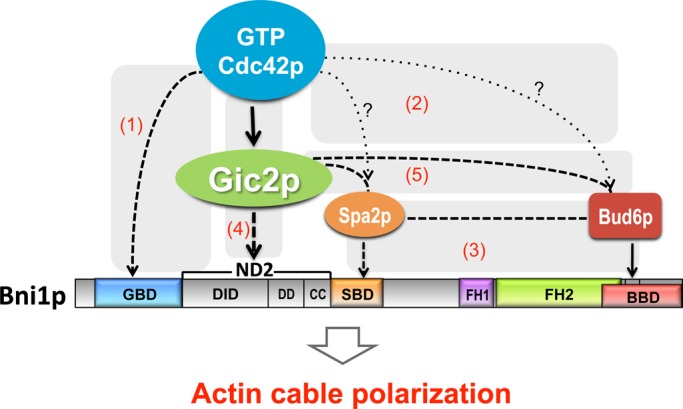
References
-
- Alberts AS. Identification of a carboxyl-terminal diaphanous-related formin homology protein autoregulatory domain. J Biol Chem. 2001;276:2824–2830. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
