

Gene flow and biological conflict systems in the origin and evolution of eukaryotes
- PMID: 22919680
- PMCID: PMC3417536
- DOI: 10.3389/fcimb.2012.00089
Gene flow and biological conflict systems in the origin and evolution of eukaryotes
Abstract
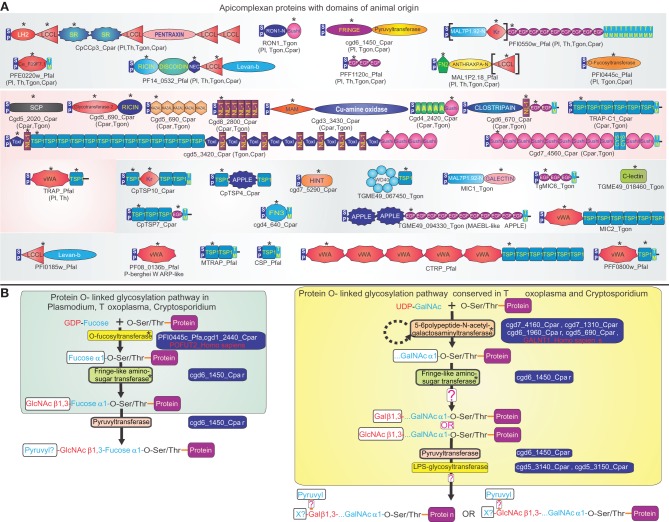
The endosymbiotic origin of eukaryotes brought together two disparate genomes in the cell. Additionally, eukaryotic natural history has included other endosymbiotic events, phagotrophic consumption of organisms, and intimate interactions with viruses and endoparasites. These phenomena facilitated large-scale lateral gene transfer and biological conflicts. We synthesize information from nearly two decades of genomics to illustrate how the interplay between lateral gene transfer and biological conflicts has impacted the emergence of new adaptations in eukaryotes. Using apicomplexans as example, we illustrate how lateral transfer from animals has contributed to unique parasite-host interfaces comprised of adhesion- and O-linked glycosylation-related domains. Adaptations, emerging due to intense selection for diversity in the molecular participants in organismal and genomic conflicts, being dispersed by lateral transfer, were subsequently exapted for eukaryote-specific innovations. We illustrate this using examples relating to eukaryotic chromatin, RNAi and RNA-processing systems, signaling pathways, apoptosis and immunity. We highlight the major contributions from catalytic domains of bacterial toxin systems to the origin of signaling enzymes (e.g., ADP-ribosylation and small molecule messenger synthesis), mutagenic enzymes for immune receptor diversification and RNA-processing. Similarly, we discuss contributions of bacterial antibiotic/siderophore synthesis systems and intra-genomic and intra-cellular selfish elements (e.g., restriction-modification, mobile elements and lysogenic phages) in the emergence of chromatin remodeling/modifying enzymes and RNA-based regulation. We develop the concept that biological conflict systems served as evolutionary "nurseries" for innovations in the protein world, which were delivered to eukaryotes via lateral gene flow to spur key evolutionary innovations all the way from nucleogenesis to lineage-specific adaptations.
Keywords: RNAi; antibiotics; biological conflict; endosymbiosis; immunity proteins; restriction-modfication; selfish elements; toxins.
Figures


References
-
- Adachi J., Hasegawa M. (1992). MOLPHY: Programs for Molecular Phylogenetics. Tokyo: Institute of Statistical Mathematics
-
- Aepfelbacher M., Aktories K., Just I. (2000). Bacterial Protein Toxins. Berlin, New York: Springer
-
- Allis C. D., Jenuwein T., Reinberg D., Caparros M. (2006). Epigenetics. New York, NY: Cold Spring Harbor Laboratory Press
-
- Alouf J. E., Popoff M. R. (2006). The Comprehensive Sourcebook of Bacterial Protein Toxins. Amsterdam, Boston: Elsevier Academic Press
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources