

Generation and characterization of non-competitive furin-inhibiting nanobodies
- PMID: 22920187
- PMCID: PMC4737579
- DOI: 10.1042/BJ20120537
Generation and characterization of non-competitive furin-inhibiting nanobodies
Abstract
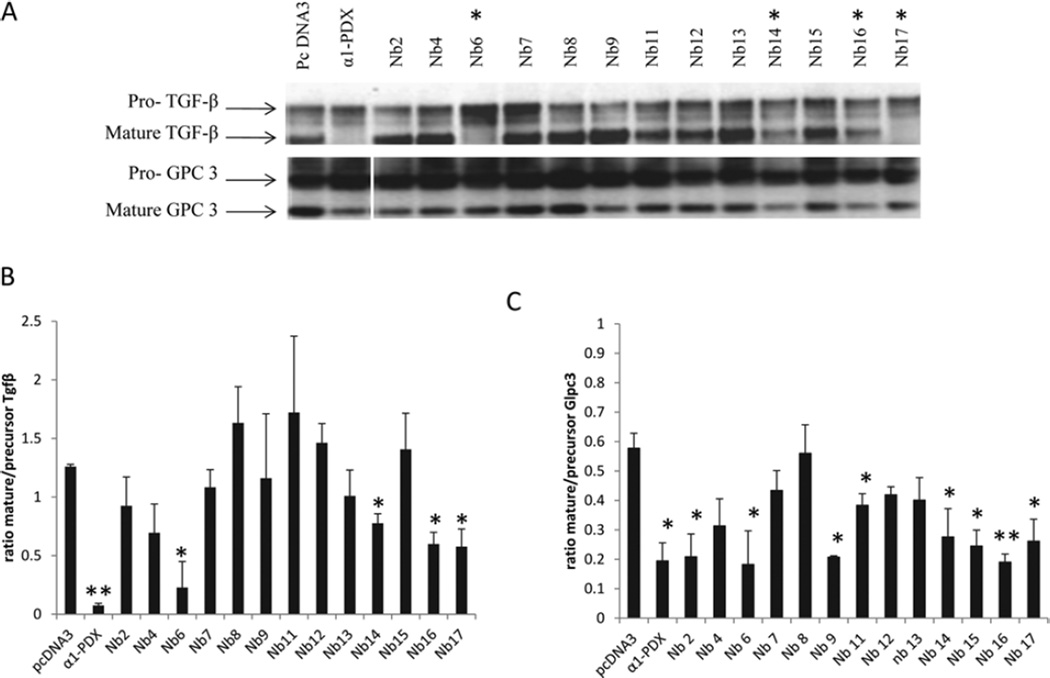
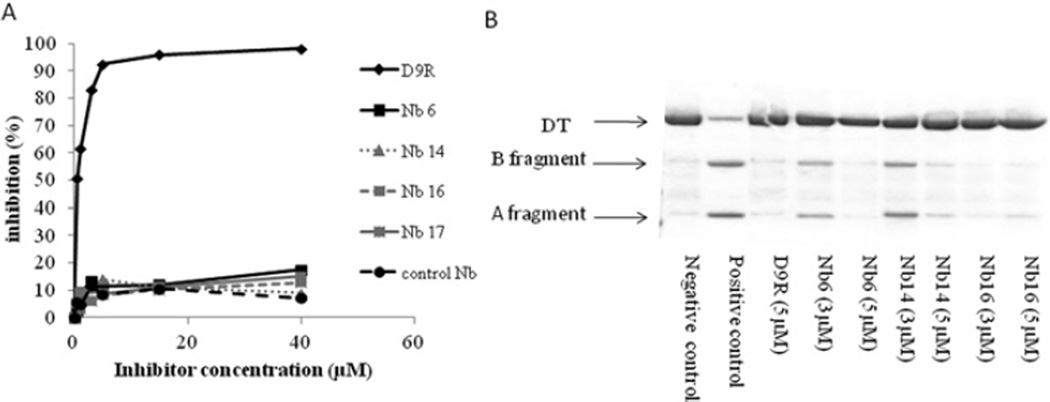
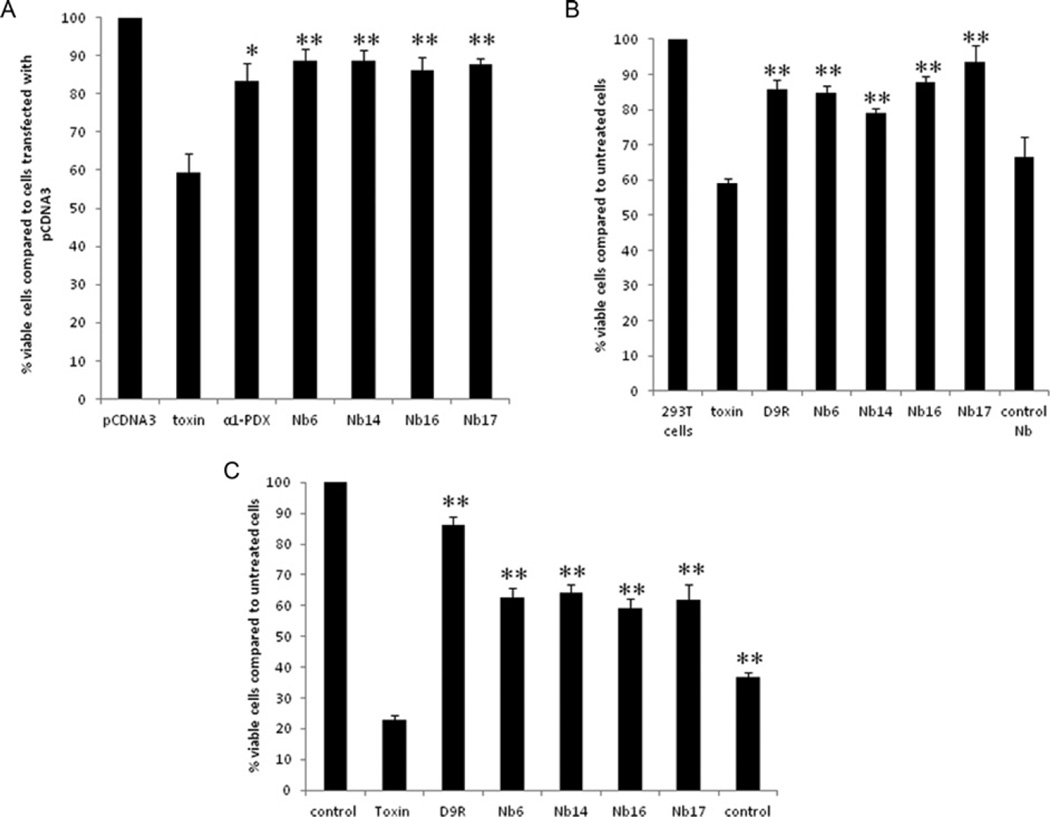
The PC (proprotein convertase) furin cleaves a large variety of proproteins and hence plays a major role in many pathologies. Therefore furin inhibition might be a good strategy for therapeutic intervention, and several furin inhibitors have been generated, although none are entirely furin-specific. To reduce potential side effects caused by cross-reactivity with other proteases, dromedary heavy-chain-derived nanobodies against catalytically active furin were developed as specific furin inhibitors. The nanobodies bound only to furin but not to other PCs. Upon overexpression in cell lines, they inhibited the cleavage of two different furin substrates, TGFβ (transforming growth factor β) and GPC3 (glypican 3). Purified nanobodies could inhibit the cleavage of diphtheria toxin into its enzymatically active A fragment, but did not inhibit cleavage of a small synthetic peptide-based substrate, suggesting a mode-of-action based on steric hindrance. The dissociation constant of purified nanobody 14 is in the nanomolar range. The nanobodies were non-competitive inhibitors with an inhibitory constant in the micromolar range as demonstrated by Dixon plot. Furthermore, anti-furin nanobodies could protect HEK (human embryonic kidney)-293T cells from diphtheria-toxin-induced cytotoxicity as efficiently as the PC inhibitor nona-D-arginine. In conclusion, these antibody-based single-domain nanobodies represent the first generation of highly specific non-competitive furin inhibitors.
Figures



References
-
- Taylor NA, Van De Ven WJ, Creemers JW. Curbing activation: proprotein convertases in homeostasis and pathology. FASEB J. 2003;17:1215–1227. - PubMed
-
- Creemers JW, Khatib AM. Knock-out mouse models of proprotein convertases: unique functions or redundancy? Front. Biosci. 2008;13:4960–4971. - PubMed
-
- Roebroek AJ, Umans L, Pauli IG, Robertson EJ, van Leuven F, Van de Ven WJ, Constam DB. Failure of ventral closure and axial rotation in embryos lacking the proprotein convertase Furin. Development. 1998;125:4863–4876. - PubMed
-
- Roebroek AJ, Taylor NA, Louagie E, Pauli I, Smeijers L, Snellinx A, Lauwers A, Van de Ven WJ, Hartmann D, Creemers JW. Limited redundancy of the proprotein convertase furin in mouse liver. J. Biol. Chem. 2004;279:53442–53450. - PubMed
-
- De Vos L, Declercq J, Rosas GG, Van Damme B, Roebroek A, Vermorken F, Ceuppens J, van de Ven W, Creemers J. MMTV-cre-mediated fur inactivation concomitant with PLAG1 proto-oncogene activation delays salivary gland tumorigenesis in mice. Int. J. Oncol. 2008;32:1073–1083. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous