

A broad range of self-reactivity drives thymic regulatory T cell selection to limit responses to self
- PMID: 22921379
- PMCID: PMC3456990
- DOI: 10.1016/j.immuni.2012.07.009
A broad range of self-reactivity drives thymic regulatory T cell selection to limit responses to self
Abstract
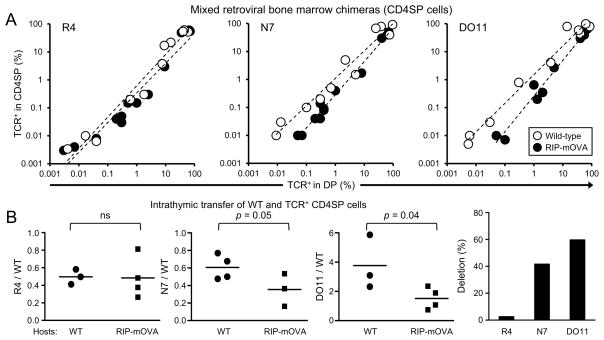
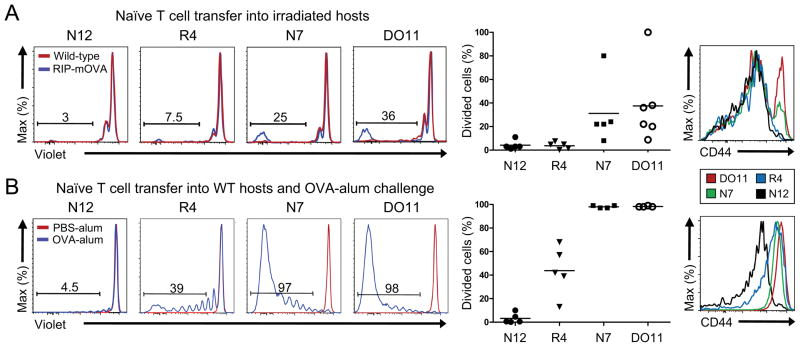
The degree of T cell self-reactivity considered dangerous by the immune system, thereby requiring thymic selection processes to prevent autoimmunity, is unknown. Here, we analyzed a panel of T cell receptors (TCRs) with a broad range of reactivity to ovalbumin (OVA(323-339)) in the rat insulin promoter (RIP)-mOVA self-antigen model for their ability to trigger thymic self-tolerance mechanisms. Thymic regulatory T (Treg) cell generation in vivo was directly correlated with in vitro TCR reactivity to OVA-peptide in a broad ~1,000-fold range. Interestingly, higher TCR affinity was associated with a larger Treg cell developmental "niche" size, even though the amount of antigen should remain constant. The TCR-reactivity threshold to elicit thymic negative selection and peripheral T cell responses was ~100-fold higher than that of Treg cell differentiation. Thus, these data suggest that the broad range of self-reactivity that elicits thymic Treg cell generation is tuned to secure peripheral tolerance to self.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures




Comment in
-
Regulatory T cell differentiation: turning harmful into useful.Immunity. 2012 Sep 21;37(3):441-3. doi: 10.1016/j.immuni.2012.09.002. Immunity. 2012. PMID: 22999949
References
-
- Apostolou I, Sarukhan A, Klein L, von Boehmer H. Origin of regulatory T cells with known specificity for antigen. Nature Immunology. 2002;3:756–763. - PubMed
-
- Bouneaud C, Kourilsky P, Bousso P. Impact of negative selection on the T cell repertoire reactive to a self-peptide: a large fraction of T cell clones escapes clonal deletion. Immunity. 2000;13:829–840. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
