

A bistable circuit involving SCARECROW-RETINOBLASTOMA integrates cues to inform asymmetric stem cell division
- PMID: 22921914
- PMCID: PMC3500399
- DOI: 10.1016/j.cell.2012.07.017
A bistable circuit involving SCARECROW-RETINOBLASTOMA integrates cues to inform asymmetric stem cell division
Abstract
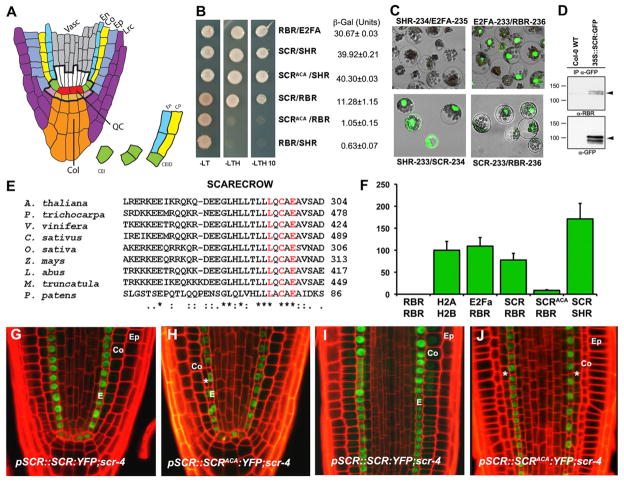
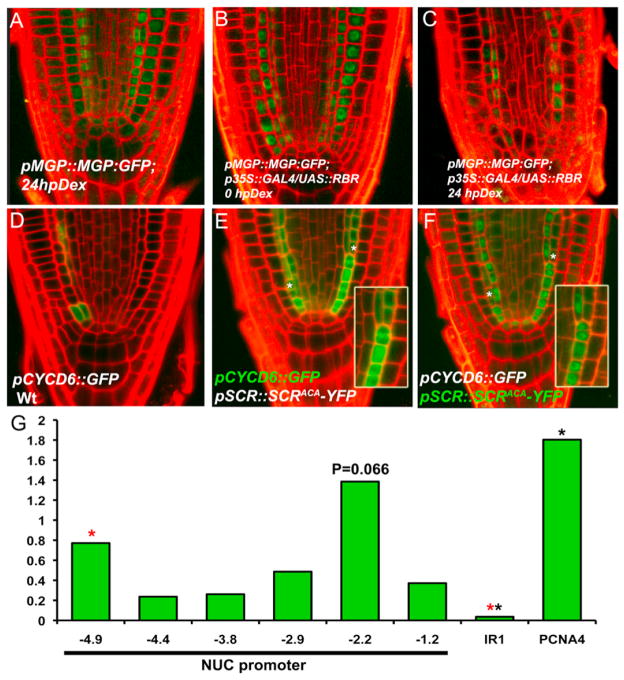
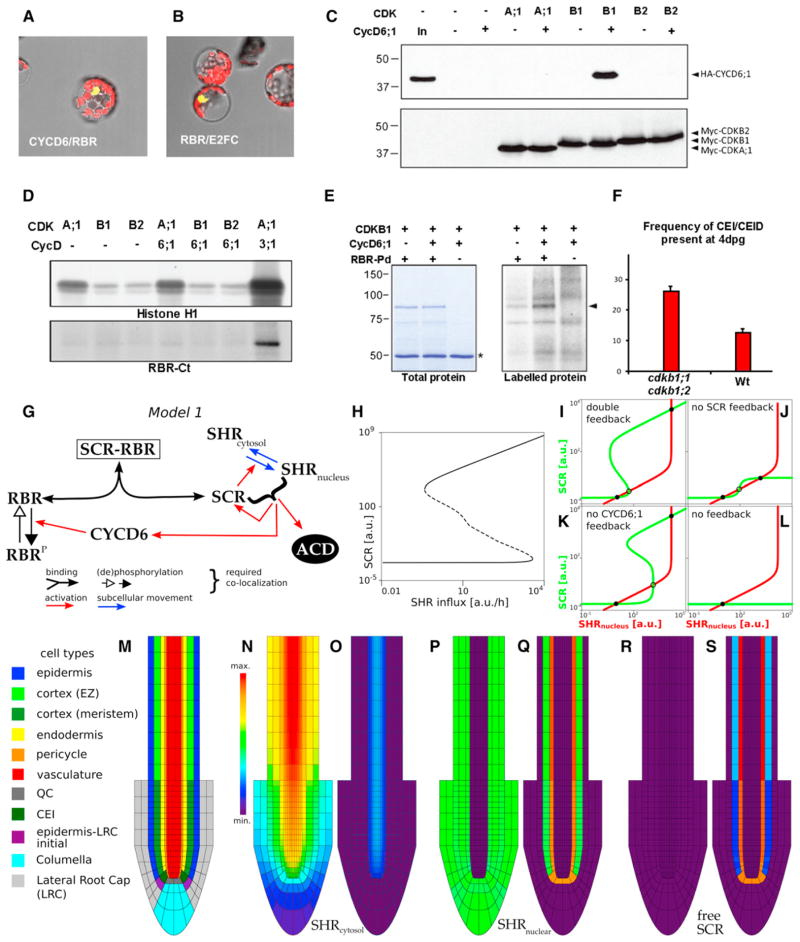
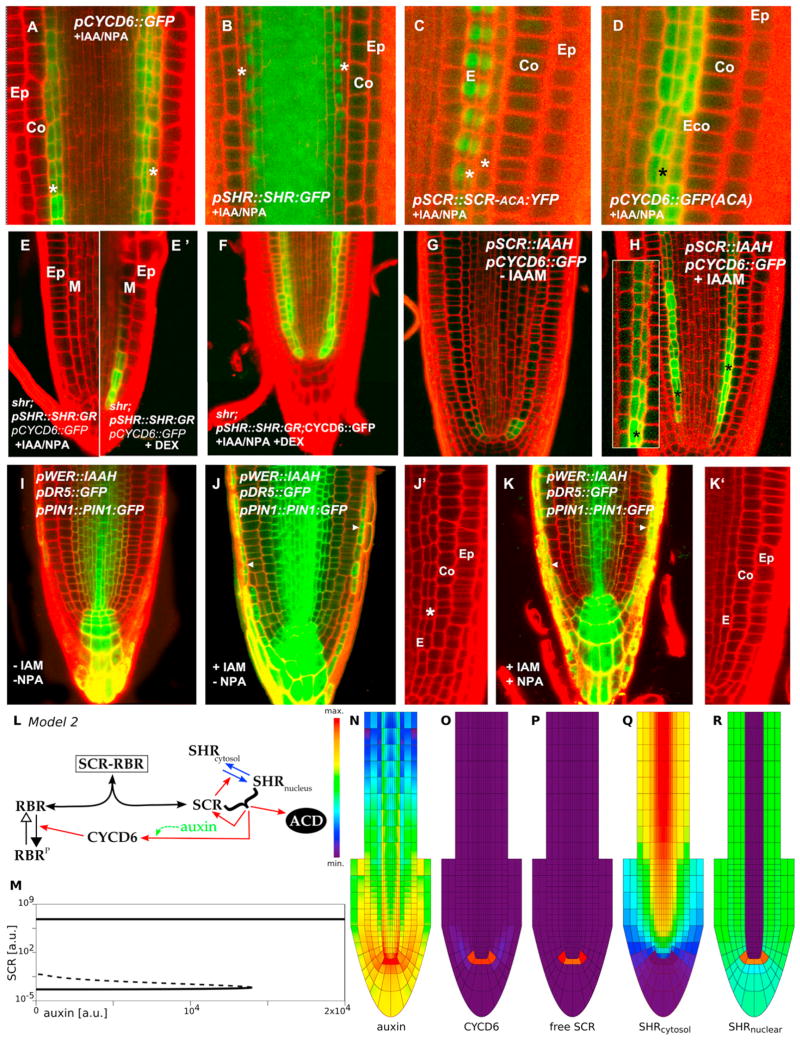
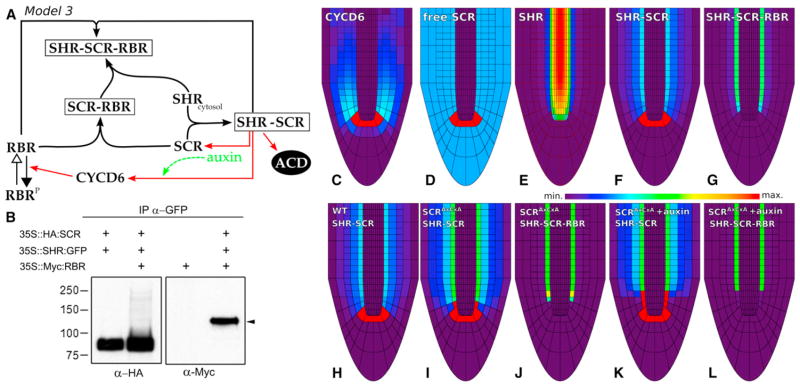
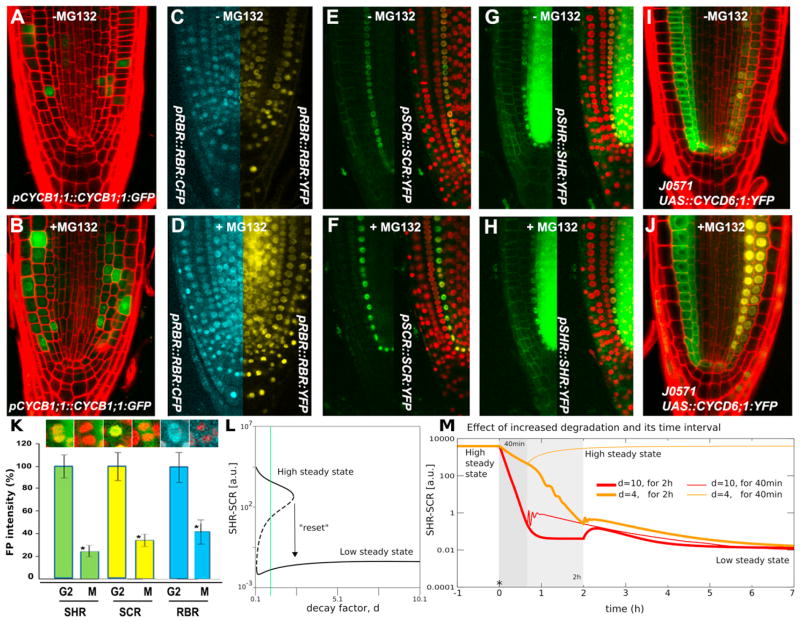
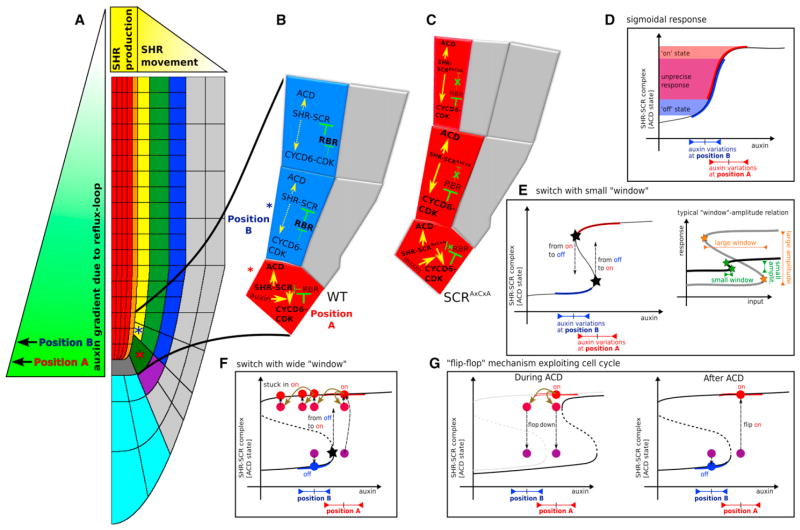
In plants, where cells cannot migrate, asymmetric cell divisions (ACDs) must be confined to the appropriate spatial context. We investigate tissue-generating asymmetric divisions in a stem cell daughter within the Arabidopsis root. Spatial restriction of these divisions requires physical binding of the stem cell regulator SCARECROW (SCR) by the RETINOBLASTOMA-RELATED (RBR) protein. In the stem cell niche, SCR activity is counteracted by phosphorylation of RBR through a cyclinD6;1-CDK complex. This cyclin is itself under transcriptional control of SCR and its partner SHORT ROOT (SHR), creating a robust bistable circuit with either high or low SHR-SCR complex activity. Auxin biases this circuit by promoting CYCD6;1 transcription. Mathematical modeling shows that ACDs are only switched on after integration of radial and longitudinal information, determined by SHR and auxin distribution, respectively. Coupling of cell-cycle progression to protein degradation resets the circuit, resulting in a "flip flop" that constrains asymmetric cell division to the stem cell region.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Abrash EB, Bergmann DC. Asymmetric cell divisions: a view from plant development. Dev Cell. 2009;16:783–796. - PubMed
-
- Alon U. Network motifs: theory and experimental approaches. Nat Rev Genet. 2007;8:450–461. - PubMed
-
- Boniotti MB, Gutierrez C. A cell-cycle-regulated kinase activity phosphorylates plant retinoblastoma protein and contains, in Arabidopsis, a CDKA/cyclin D complex. Plant J. 2001;28:341–350. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/E022383/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/J/000C0646/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- R01 GM043778/GM/NIGMS NIH HHS/United States
- BB/J004553/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/G00482X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/G00482X/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E022383/2/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- R01-GM043778/GM/NIGMS NIH HHS/United States
- P50 GM081883/GM/NIGMS NIH HHS/United States
- BBS/E/J/000C0645/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
