

Progestin modulates the lipid profile and sensitivity of breast cancer cells to docetaxel
- PMID: 22922095
- PMCID: PMC4671297
- DOI: 10.1016/j.mce.2012.08.005
Progestin modulates the lipid profile and sensitivity of breast cancer cells to docetaxel
Abstract
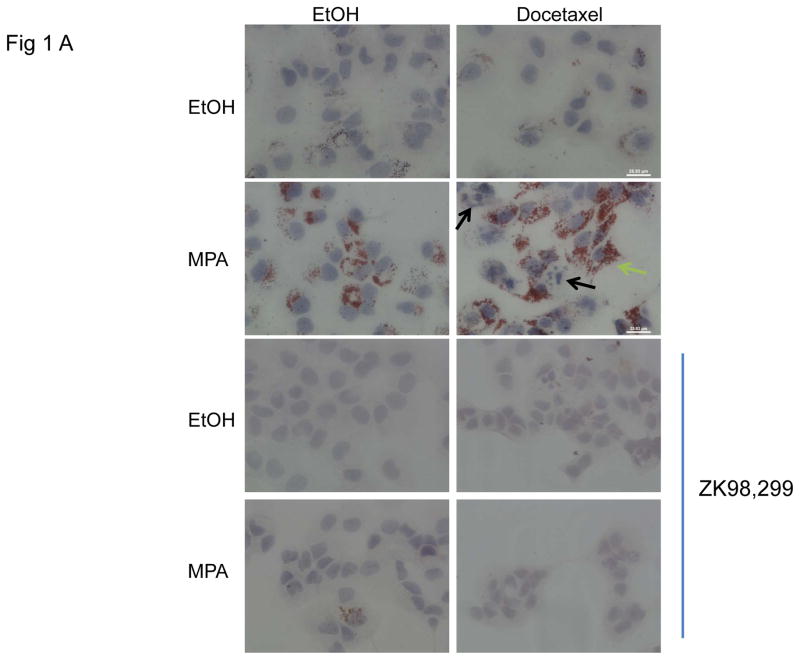
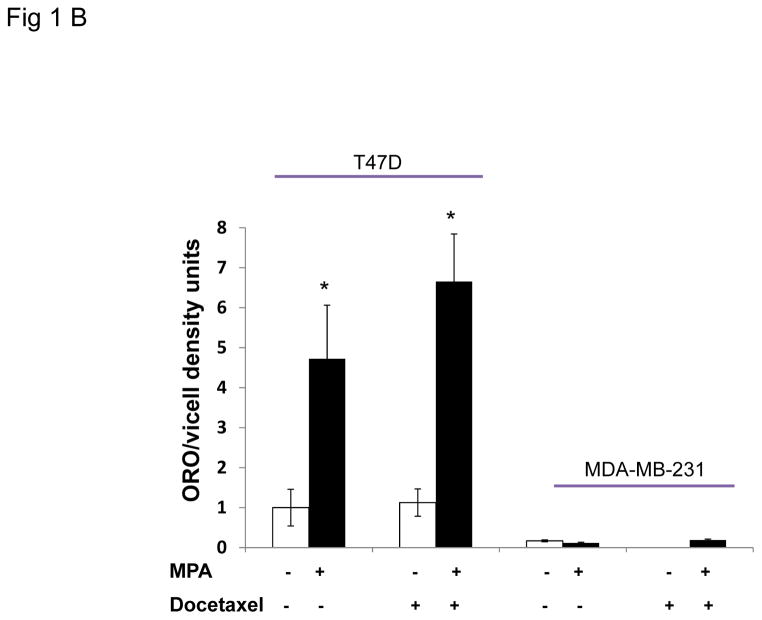
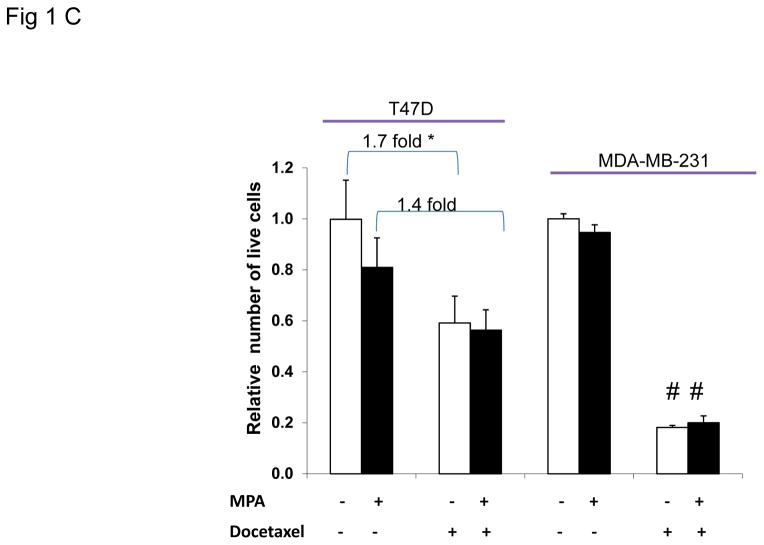
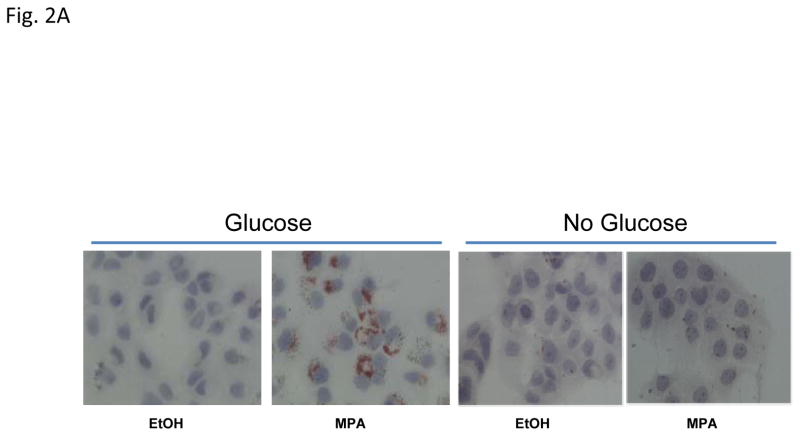
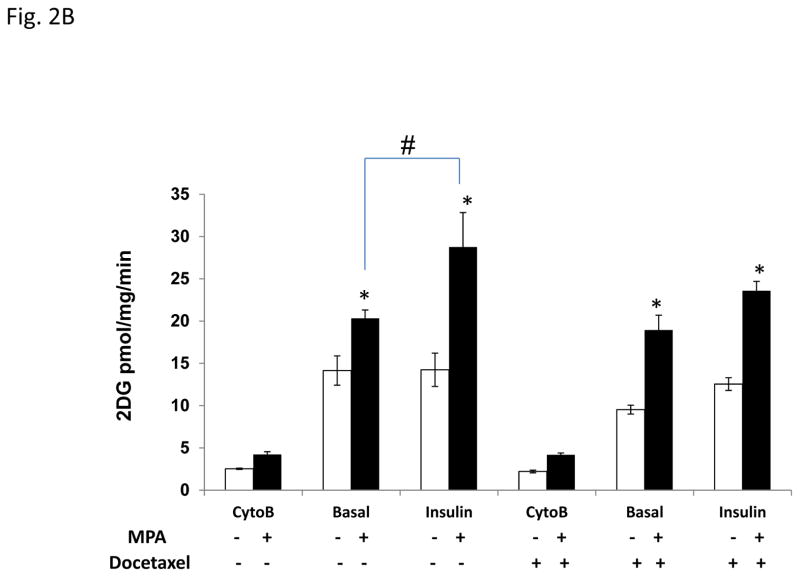
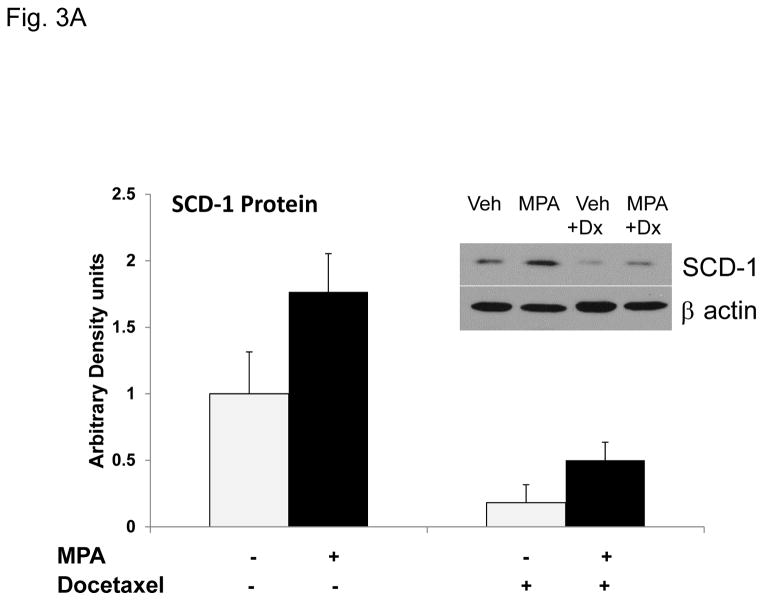
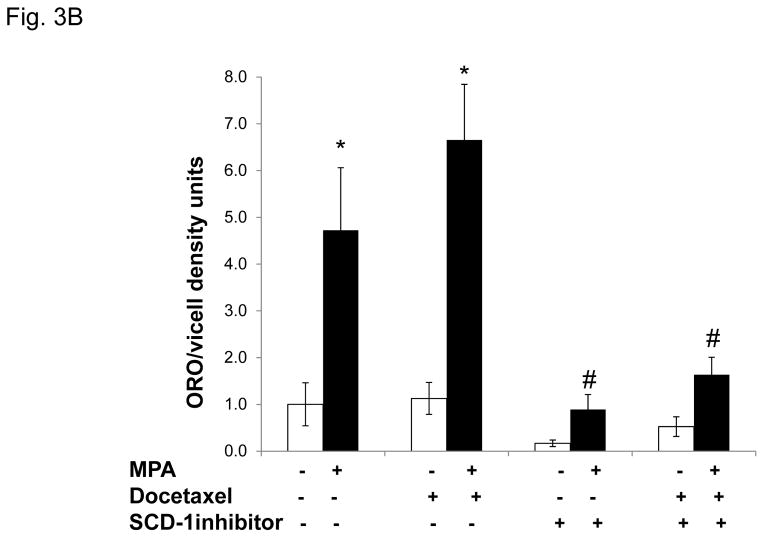
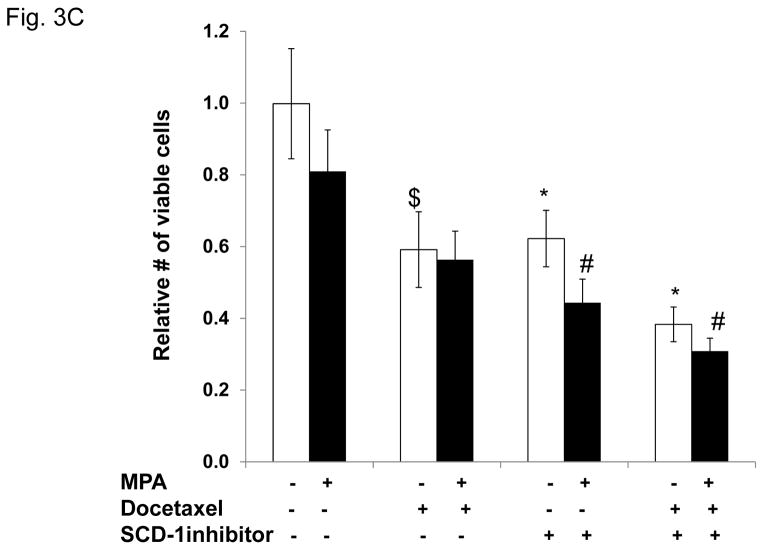
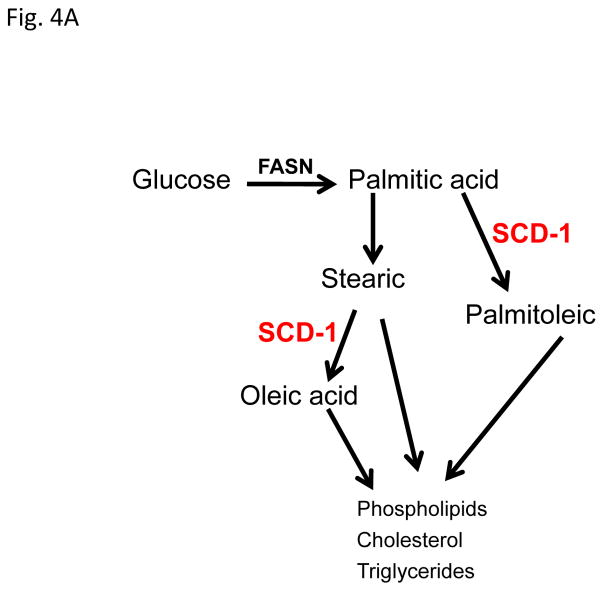
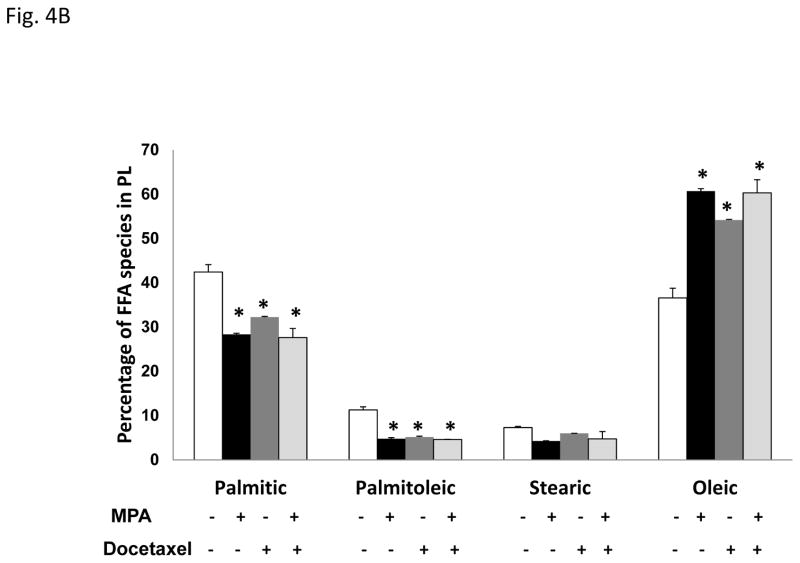
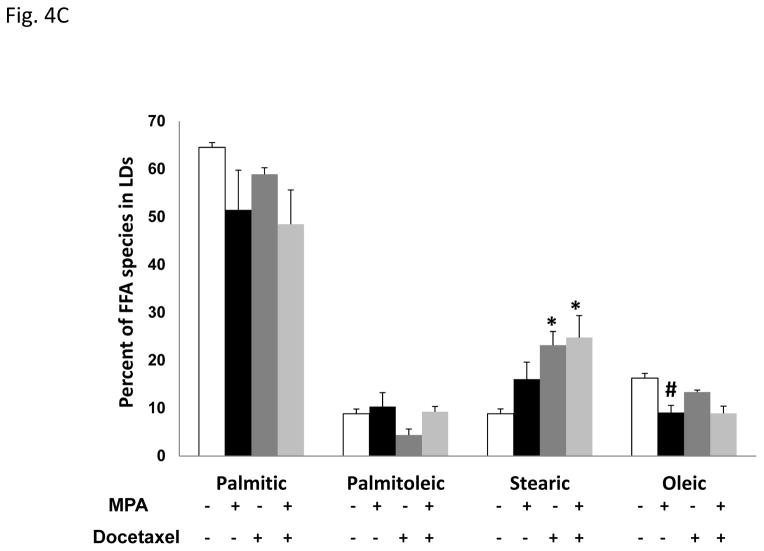
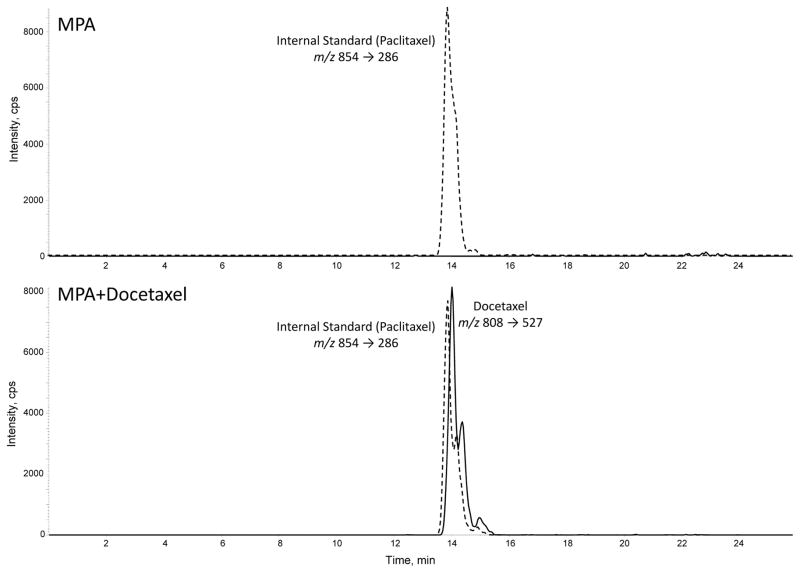
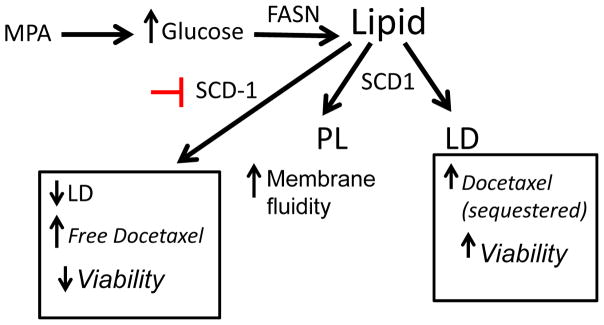
Progestins induce lipid accumulation in progesterone receptor (PR)-positive breast cancer cells. We speculated that progestin-induced alterations in lipid biology confer resistance to chemotherapy. To examine the biology of lipid loaded breast cancer cells, we used a model of progestin-induced lipid synthesis. T47D (PR-positive) and MDA-MB-231 (PR-negative) cell lines were used to study progestin response. Oil red O staining of T47D cells treated with progestin showed lipid droplet formation was PR dependent, glucose dependent and reduced sensitivity to docetaxel. This protection was not observed in PR-negative MDA-MB-231 cells. Progestin treatment induced stearoyl CoA desaturase-1 (SCD-1) enzyme expression and chemical inhibition of SCD-1 diminished lipid droplets and cell viability, suggesting the importance of lipid stores in cancer cell survival. Gas chromatography/mass spectroscopy analysis of phospholipids from progestin-treated T47D cells revealed an increase in unsaturated fatty acids, with oleic acid as most abundant. Cells surviving docetaxel treatment also contained more oleic acid in phospholipids, suggesting altered membrane fluidity as a potential mechanism of chemoresistance mediated in part by SCD-1. Lastly, intact docetaxel molecules were present within progestin induced lipid droplets, suggesting a protective quenching effect of intracellular lipid droplets. Our studies suggest the metabolic adaptations produced by progestin provide novel metabolic targets for future combinatorial therapies for progestin-responsive breast cancers.
Copyright © 2012 Elsevier Ireland Ltd. All rights reserved.
Figures
References
-
- Baritaki S, Apostolakis S, Kanellou P, Dimanche-Boitrel MT, Spandidos DA, Bonavida B. Reversal of tumor resistance to apoptotic stimuli by alteration of membrane fluidity: therapeutic implications. Adv Cancer Res. 2007;98:149–190. - PubMed
-
- Bayet-Robert M, Morvan D, Chollet P, Barthomeuf C. Pharmacometabolomics of docetaxel-treated human MCF7 breast cancer cells provides evidence of varying cellular responses at high and low doses. Breast Cancer Res Treat 2009 - PubMed
-
- BLIGH EG, DYER WJ. A rapid method of total lipid extraction and purification. Can J Biochem Physiol. 1959;37:911–917. - PubMed
-
- Chajes V, Hulten K, Van Kappel AL, Winkvist A, Kaaks R, Hallmans G, Lenner P, Riboli E. Fatty-acid composition in serum phospholipids and risk of breast cancer: an incident case-control study in Sweden. Int J Cancer. 1999;83:585–590. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
