

Transcription factor Foxp3 and its protein partners form a complex regulatory network
- PMID: 22922362
- PMCID: PMC3448012
- DOI: 10.1038/ni.2402
Transcription factor Foxp3 and its protein partners form a complex regulatory network
Abstract
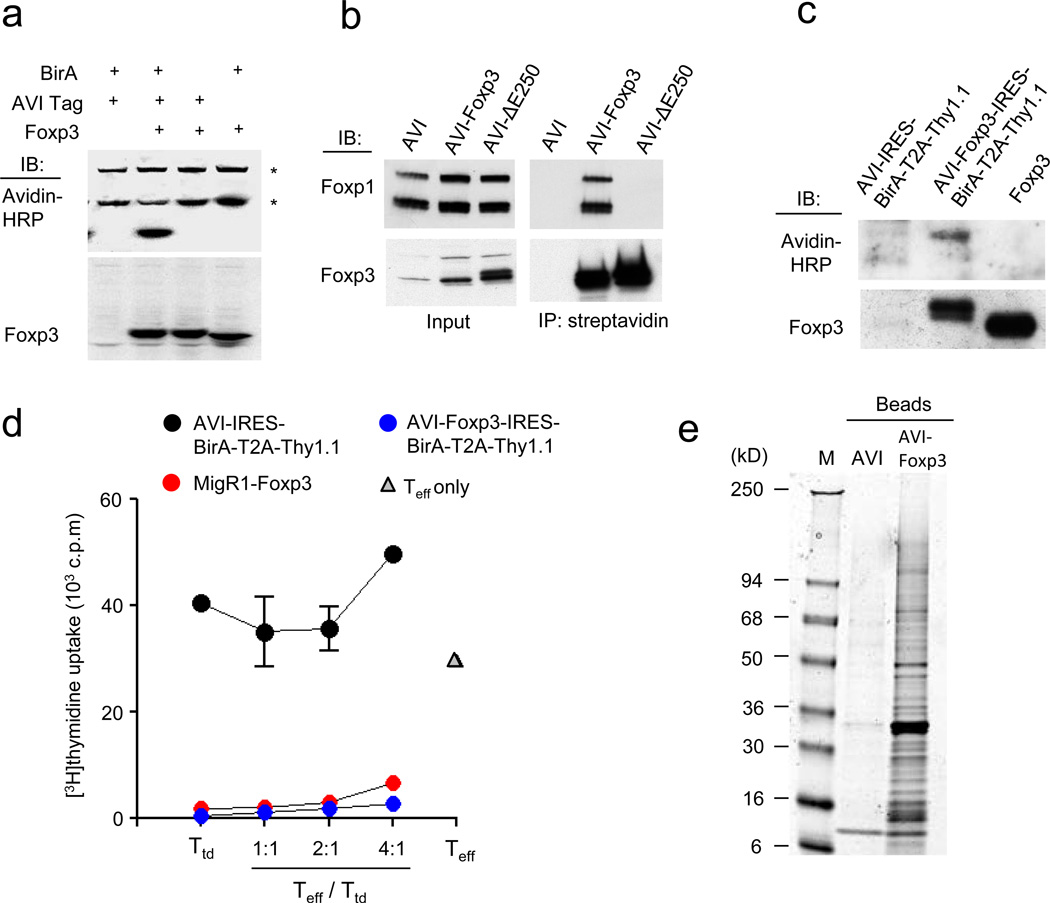
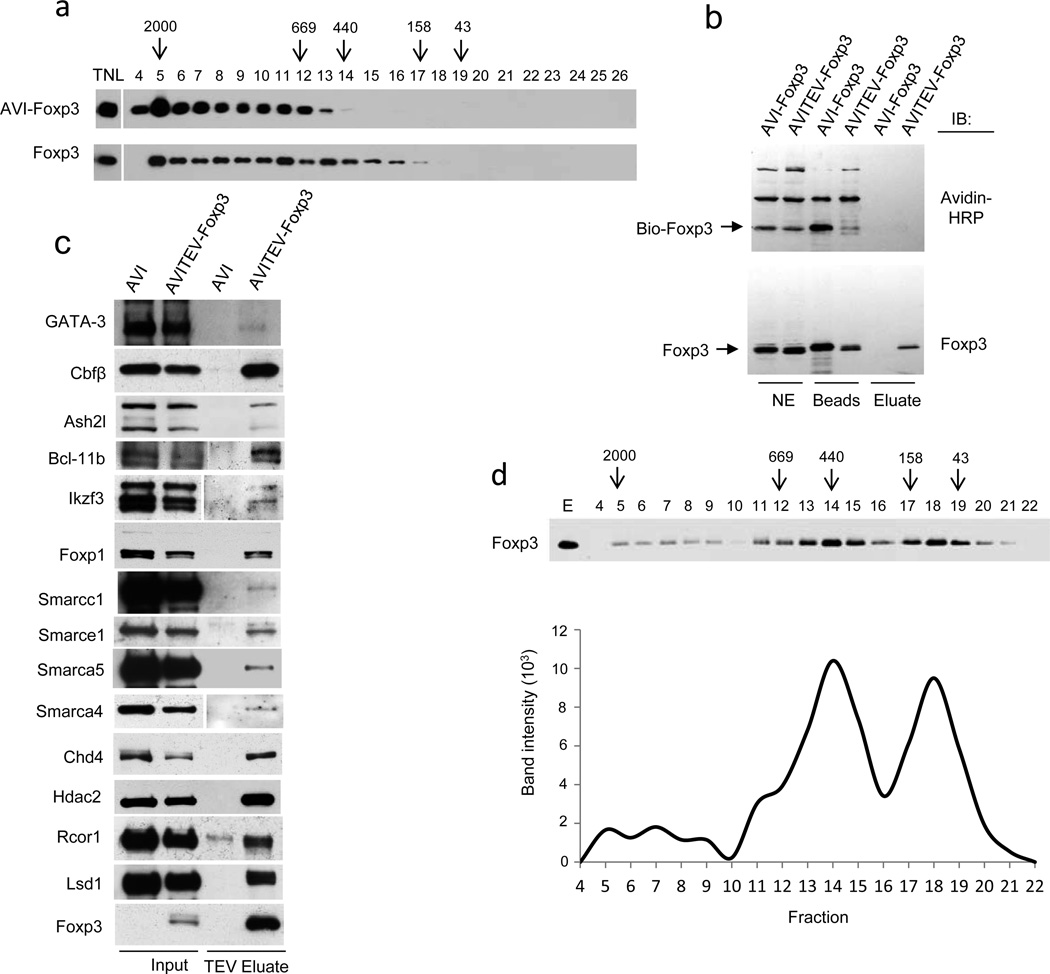
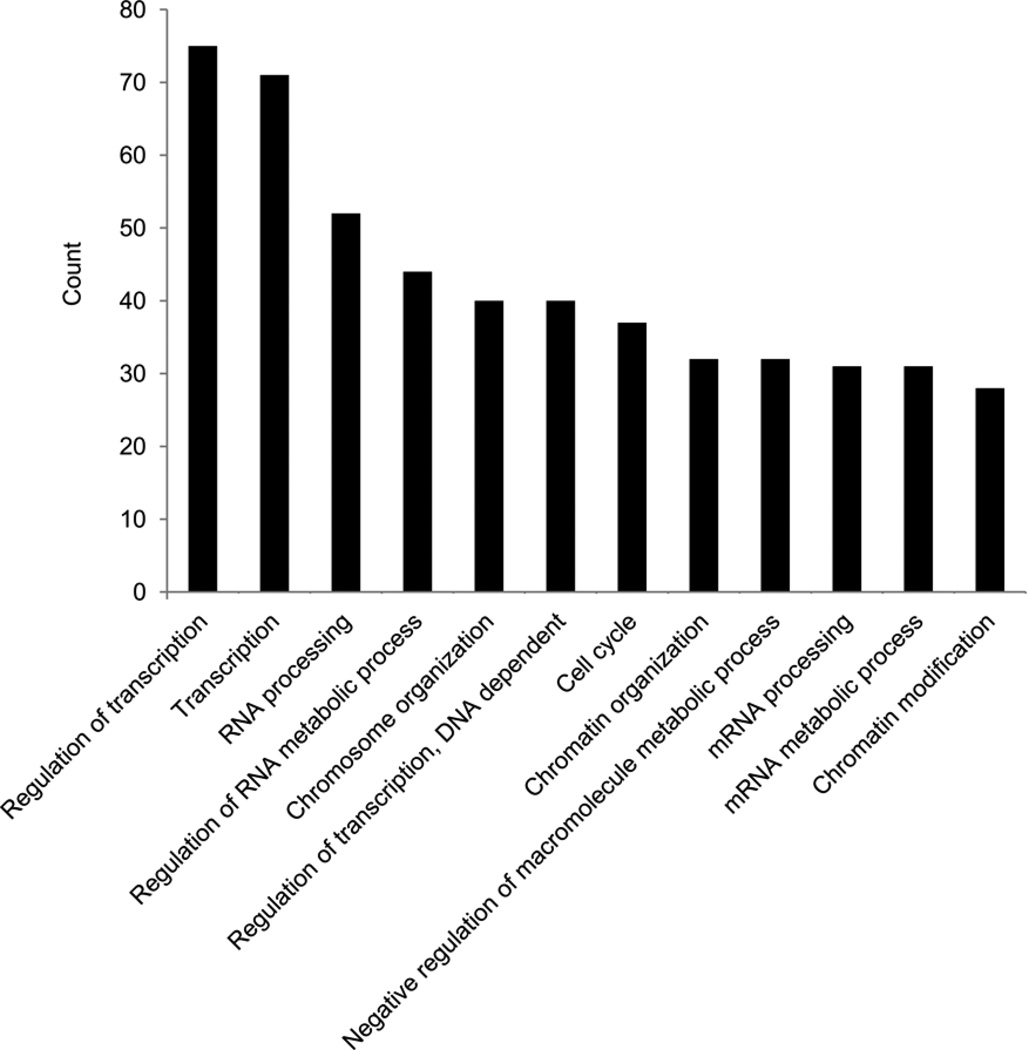
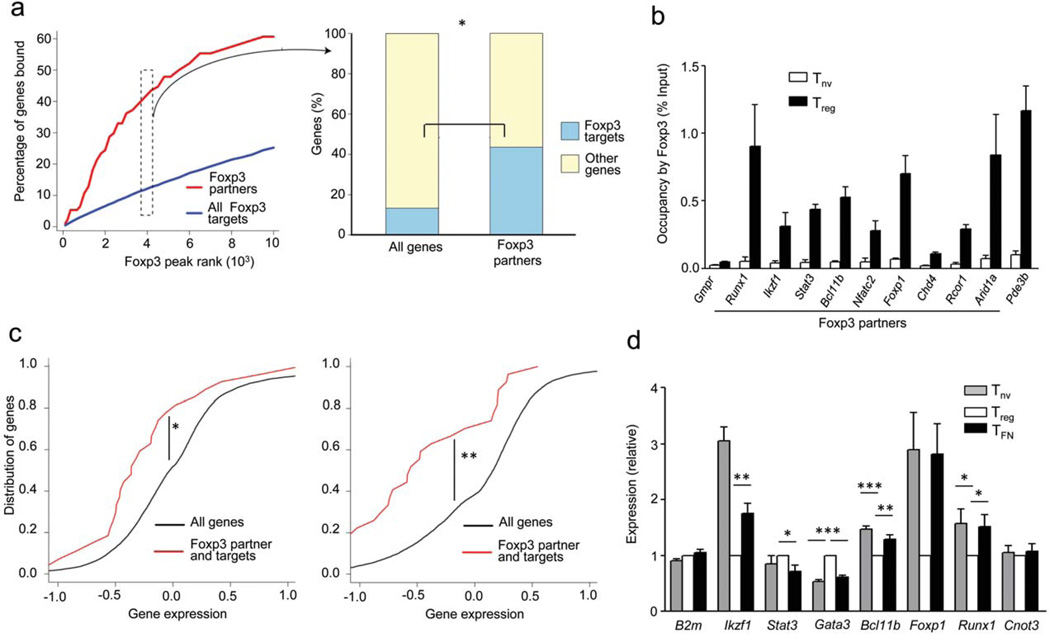
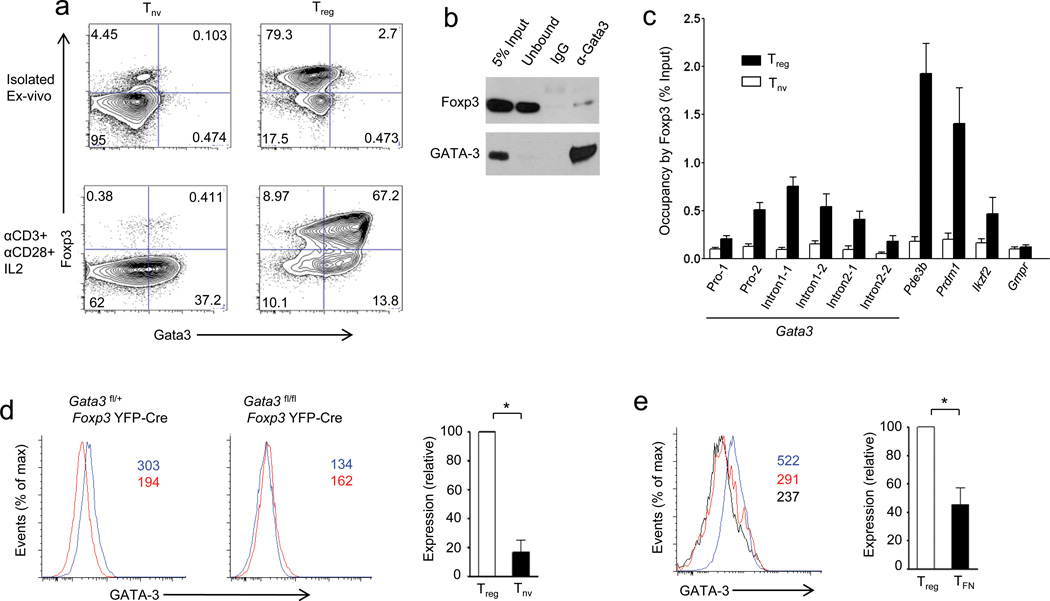
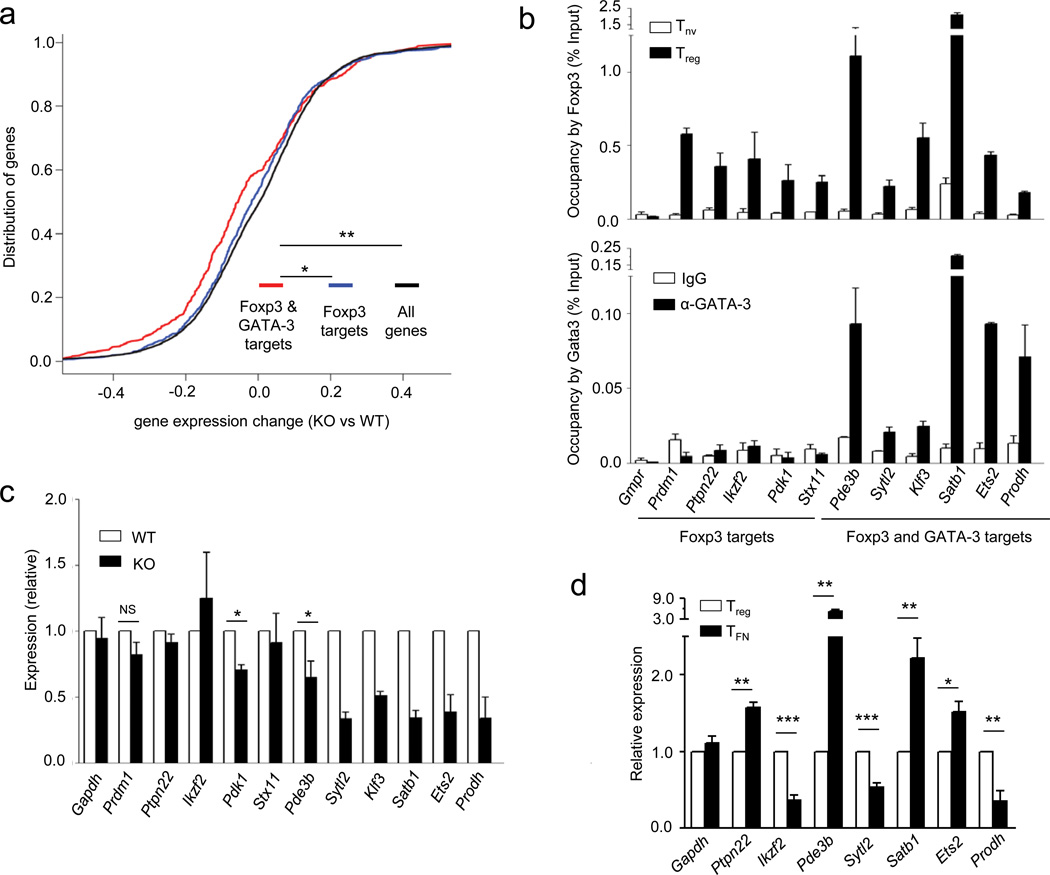
The transcription factor Foxp3 is indispensible for the differentiation and function of regulatory T cells (T(reg) cells). To gain insights into the molecular mechanisms of Foxp3-mediated gene expression, we purified Foxp3 complexes and explored their composition. Biochemical and mass-spectrometric analyses revealed that Foxp3 forms multiprotein complexes of 400-800 kDa or larger and identified 361 associated proteins, ∼30% of which were transcription related. Foxp3 directly regulated expression of a large proportion of the genes encoding its cofactors. Some transcription factor partners of Foxp3 facilitated its expression. Functional analysis of the cooperation of Foxp3 with one such partner, GATA-3, provided additional evidence for a network of transcriptional regulation afforded by Foxp3 and its associates to control distinct aspects of T(reg) cell biology.
Figures
Comment in
-
The Foxp3 interactome: a network perspective of T(reg) cells.Nat Immunol. 2012 Oct;13(10):943-5. doi: 10.1038/ni.2424. Nat Immunol. 2012. PMID: 22990900 No abstract available.
References
-
- Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and immune tolerance. Cell. 2008;133:775–787. - PubMed
-
- Bennett CL, et al. The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nat Genet. 2001;27:20–21. - PubMed
-
- Wildin RS, Freitas A. IPEX and FOXP3: clinical and research perspectives. J Autoimmun. 2005;25(Suppl):56–62. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
