

Polyethyleneimine is a potent mucosal adjuvant for viral glycoprotein antigens
- PMID: 22922673
- PMCID: PMC3496939
- DOI: 10.1038/nbt.2344
Polyethyleneimine is a potent mucosal adjuvant for viral glycoprotein antigens
Abstract
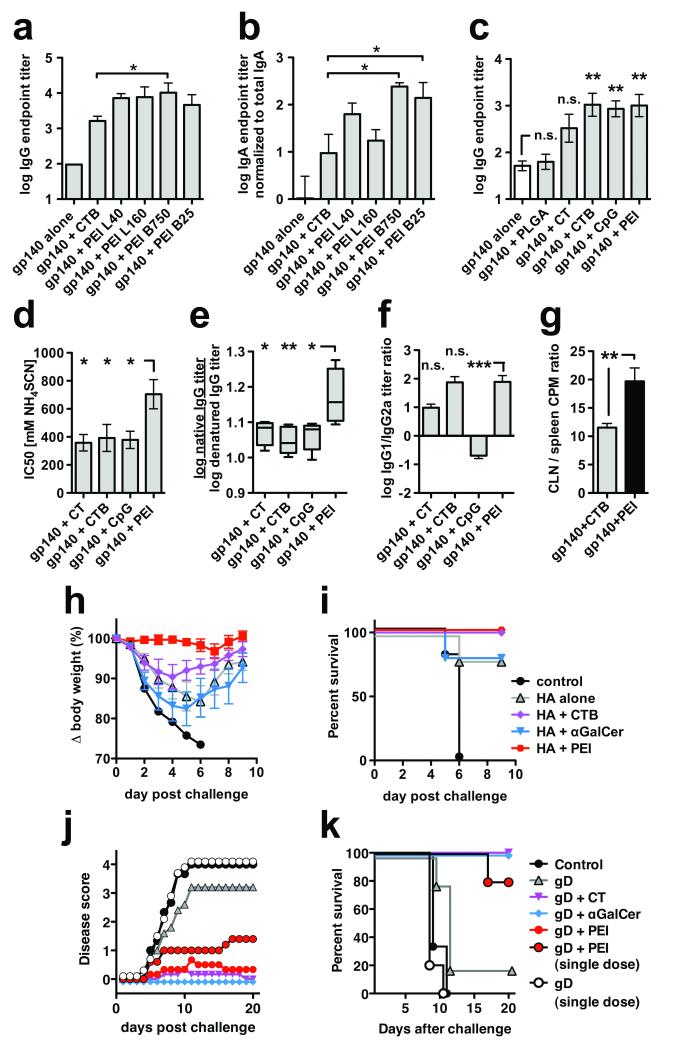
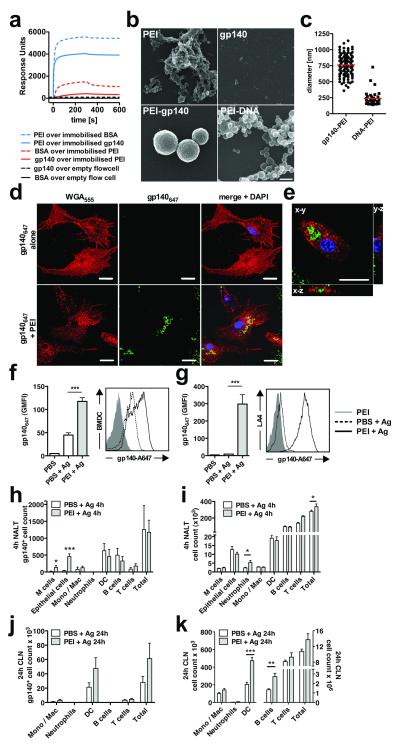
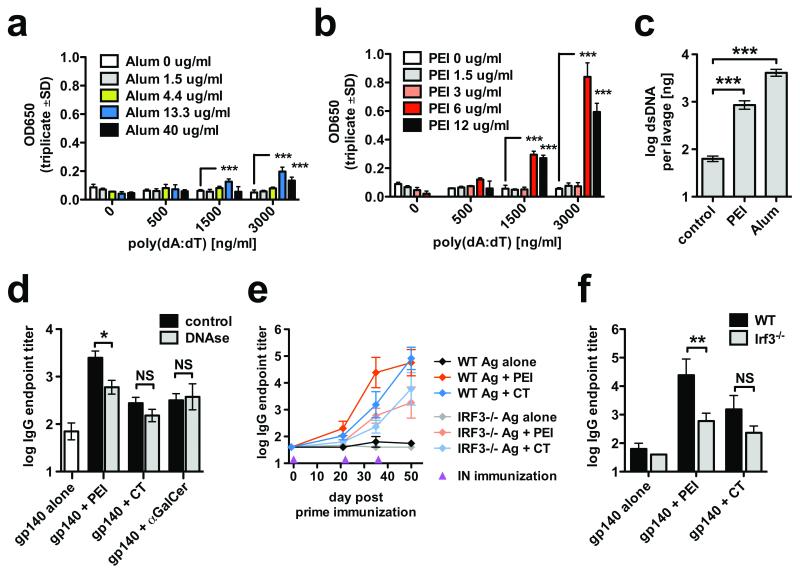
Protection against mucosally transmitted infections probably requires immunity at the site of pathogen entry, yet there are no mucosal adjuvant formulations licensed for human use. Polyethyleneimine (PEI) represents a family of organic polycations used as nucleic acid transfection reagents in vitro and DNA vaccine delivery vehicles in vivo. Here we show that diverse PEI forms have potent mucosal adjuvant activity for viral subunit glycoprotein antigens. A single intranasal administration of influenza hemagglutinin or herpes simplex virus type-2 (HSV-2) glycoprotein D with PEI elicited robust antibody-mediated protection from an otherwise lethal infection, and was superior to existing experimental mucosal adjuvants. PEI formed nanoscale complexes with antigen, which were taken up by antigen-presenting cells in vitro and in vivo, promoted dendritic cell trafficking to draining lymph nodes and induced non-proinflammatory cytokine responses. PEI adjuvanticity required release of host double-stranded DNA that triggered Irf3-dependent signaling. PEI therefore merits further investigation as a mucosal adjuvant for human use.
Figures
References
-
- Gunther M, et al. Polyethylenimines for RNAi-mediated gene targeting in vivo and siRNA delivery to the lung. Eur J Pharm Biopharm. 2010:18. - PubMed
-
- Lungwitz U, Breunig M, Blunk T, Gopferich A. Polyethylenimine-based non-viral gene delivery systems. Eur J Pharm Biopharm. 2005;60:247–266. - PubMed
-
- Fujihashi K, Koga T, van Ginkel FW, Hagiwara Y, McGhee JR. A dilemma for mucosal vaccination: efficacy versus toxicity using enterotoxin-based adjuvants. Vaccine. 2002;20:2431–2438. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
