

STAT3 signal transduction through interleukin-22 in oral squamous cell carcinoma
- PMID: 22922995
- PMCID: PMC3583669
- DOI: 10.3892/ijo.2012.1594
STAT3 signal transduction through interleukin-22 in oral squamous cell carcinoma
Abstract
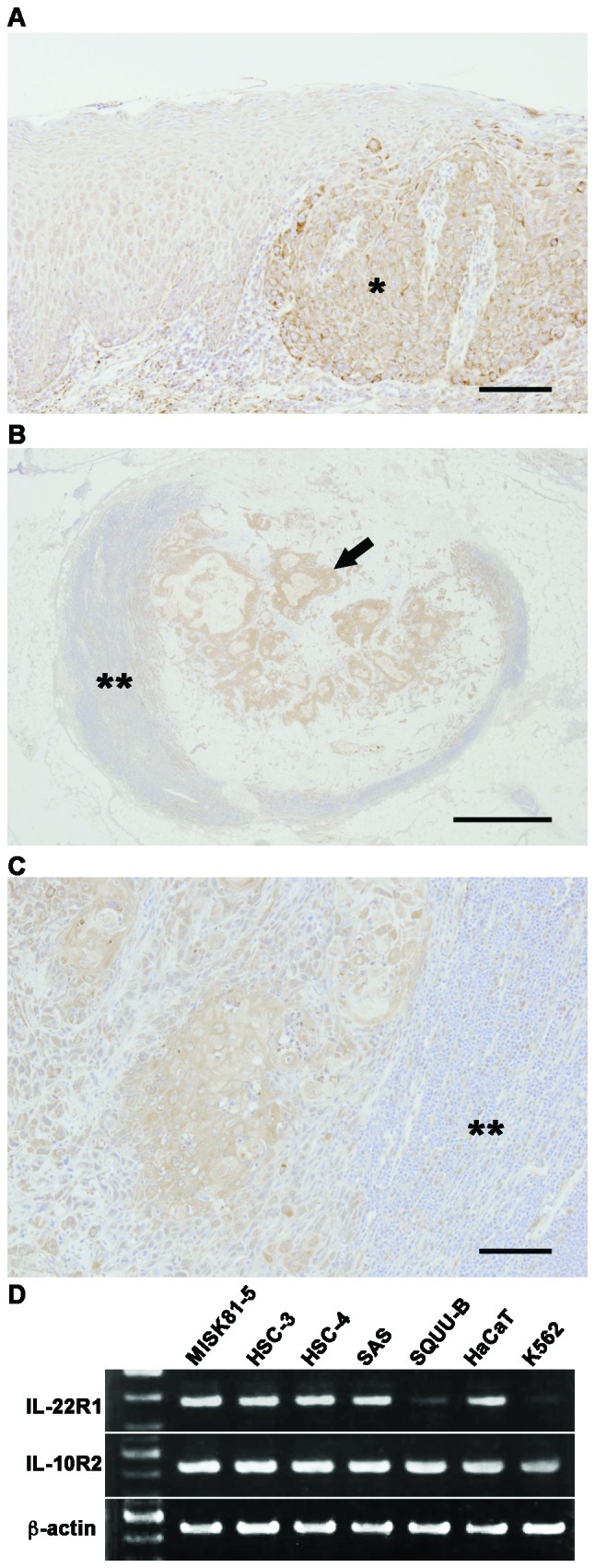
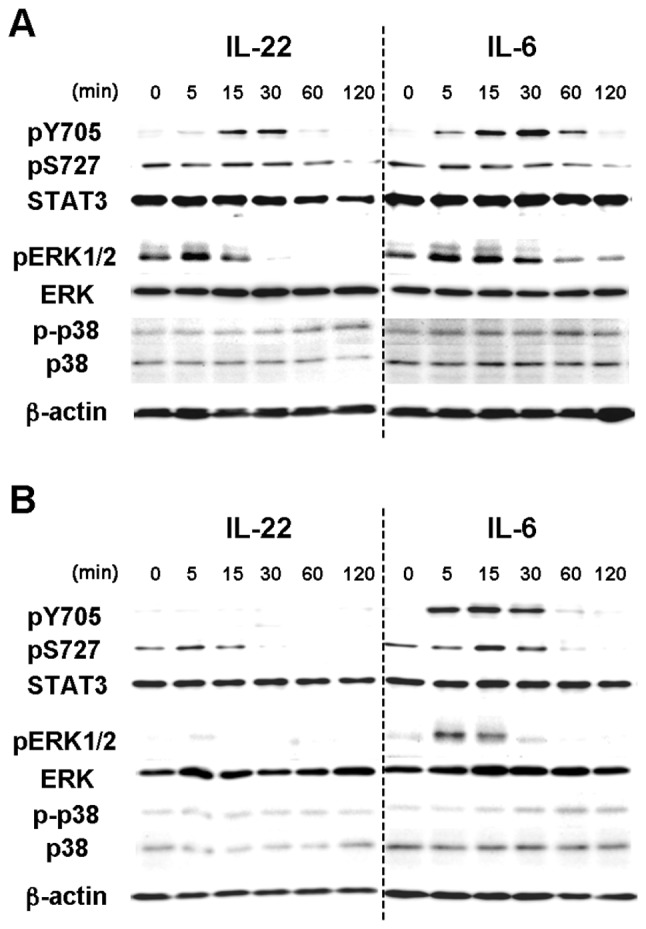
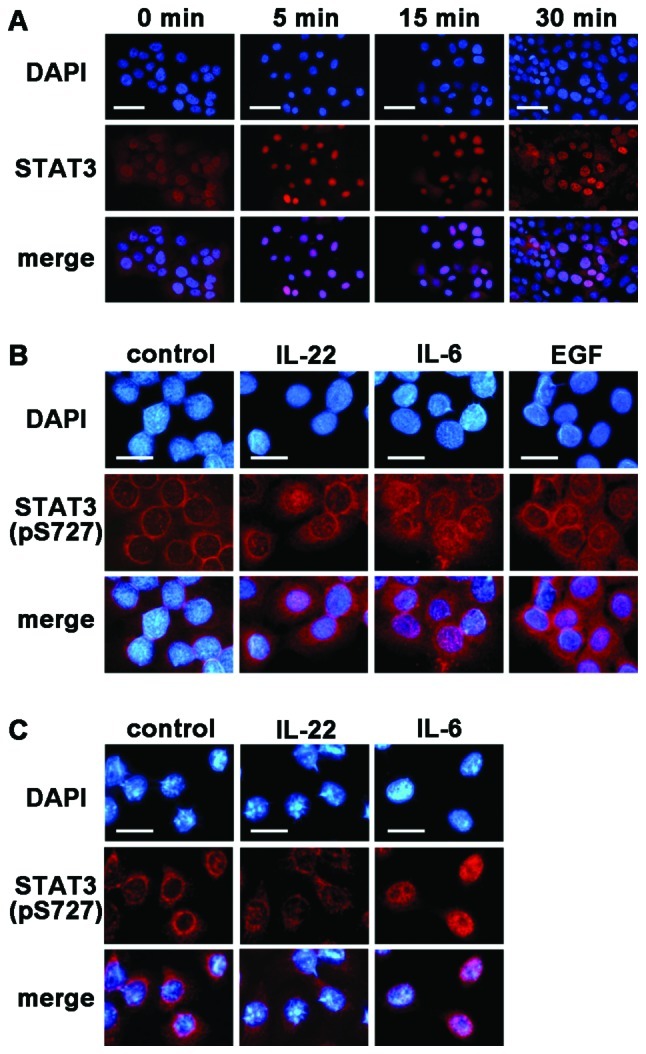
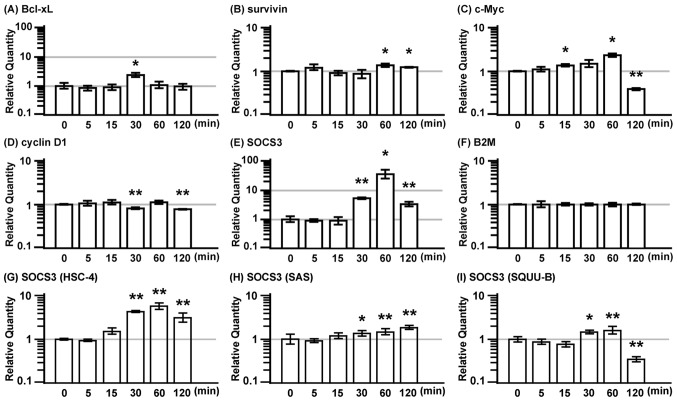
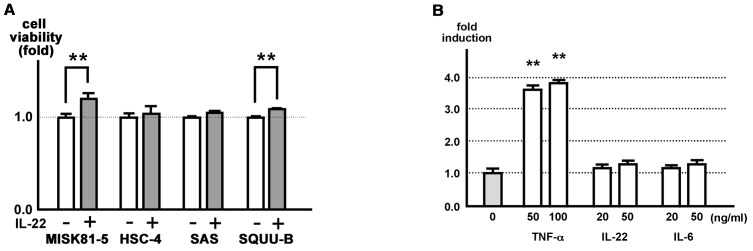
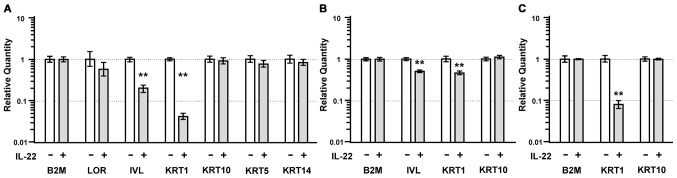
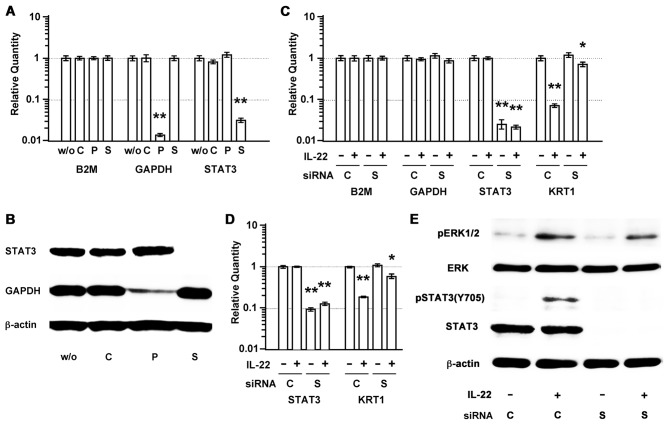
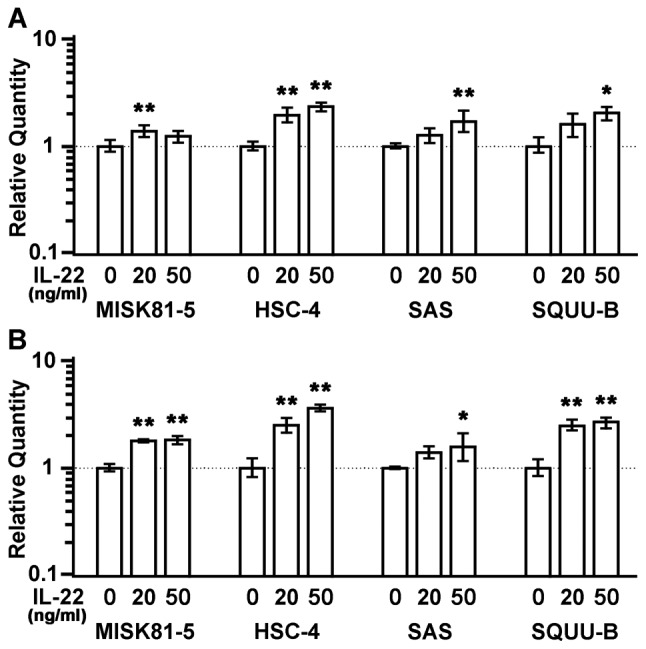
Interleukin (IL)-22 is a member of the IL-10 family. Its main targets are epithelial cells, not immune cells. We examined IL-22 signal transduction in oral squamous cell carcinoma (OSCC) cells. Immunohistochemical staining revealed that IL-22R was expressed more highly in OSCC compared to normal regions. An IL-22R signal was also observed in metastatic OSCC cells in the lymph node. RT-PCR showed that the human OSCC cell lines MISK81-5, HSC-3, HSC-4, SAS and SQUU-B expressed IL-22 receptor chains. Immunoblotting showed that IL-22 induced a transient tyrosine phosphorylation of STAT3 (pY705-STAT3) in MISK81-5 cells. The change in the serine phosphorylation of STAT3 was subtle during the examination periods. Simultaneously, pY705-STAT3 activation in HSC-3 cells was undetectable after IL-22 stimulation. Immunocytochemistry demonstrated that IL-22 induced the translocation of phosphorylated STAT3 into the nucleus of MISK81-5 cells. IL-22 temporarily upregulated the expression of anti-apoptotic and mitogenic genes such as Bcl-x, survivin and c-Myc, as well as SOCS3. IL-22 transiently activated ERK1/2 and induced a delayed phosphorylation of p38 MAP kinase, but negligibly involved the activation of NF-κB in MISK81-5 cells. MISK81-5 and SQUU-B cells treated with IL-22 showed mild cellular proliferation. MISK81-5, HSC-4 and SAS cells treated with IL-22 downregulated the keratinocyte differentiation-related genes compared with unstimulated cells. Conversely, STAT3 suppression by STAT3 siRNA strongly disrupted the downregulation of these genes by IL-22, but it did not significantly affect the activation of ERK1/2 by IL-22. The OSCC cells used in this study upregulated the expression of SERPINB3/4 (SCCA1/2), well-known SCC markers, following treatment with IL-22. These results indicate that IL-22 differentially activates the STAT3 signaling system depending on the type of OSCC. IL-22 may therefore play a role in tumor growth, cell differentiation and progression through STAT3-dependent and -independent pathways.
Figures
References
-
- Chidzonga MM, Mahomva L. Squamous cell carcinoma of the oral cavity, maxillary antrum and lip in a Zimbabwean population: a descriptive epidemiological study. Oral Oncol. 2006;42:184–189. - PubMed
-
- Chidzonga MM. Oral malignant neoplasia: a survey of 428 cases in two Zimbabwean hospitals. Oral Oncol. 2006;42:177–183. - PubMed
-
- Dimery IW, Hong WK. Overview of combined modality therapies for head and neck cancer. J Natl Cancer Inst. 1993;85:95–111. - PubMed
-
- Schantz SP, Yu GP. Head and neck cancer incidence trends in young Americans, 1973–1997, with a special analysis for tongue cancer. Arch Otolaryngol Head Neck Surg. 2002;128:268–274. - PubMed
-
- Llewellyn CD, Linklater K, Bell J, Johnson NW, Warnakulasuriya KA. Squamous cell carcinoma of the oral cavity in patients aged 45 years and under: a descriptive analysis of 116 cases diagnosed in the South East of England from 1990 to 1997. Oral Oncol. 2003;39:106–114. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
