

A naturally occurring InDel variation in BraA.FLC.b (BrFLC2) associated with flowering time variation in Brassica rapa
- PMID: 22925611
- PMCID: PMC3487953
- DOI: 10.1186/1471-2229-12-151
A naturally occurring InDel variation in BraA.FLC.b (BrFLC2) associated with flowering time variation in Brassica rapa
Abstract
Background: Flowering time is an important trait in Brassica rapa crops. FLOWERING LOCUS C (FLC) is a MADS-box transcription factor that acts as a potent repressor of flowering. Expression of FLC is silenced when plants are exposed to low temperature, which activates flowering. There are four copies of FLC in B. rapa. Analyses of different segregating populations have suggested that BraA.FLC.a (BrFLC1) and BraA.FLC.b (BrFLC2) play major roles in controlling flowering time in B. rapa.
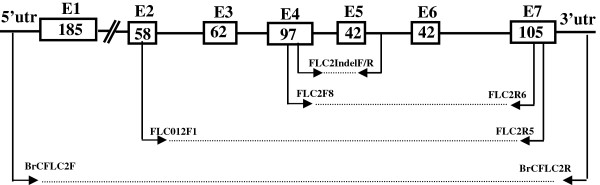
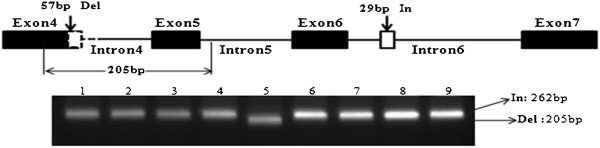
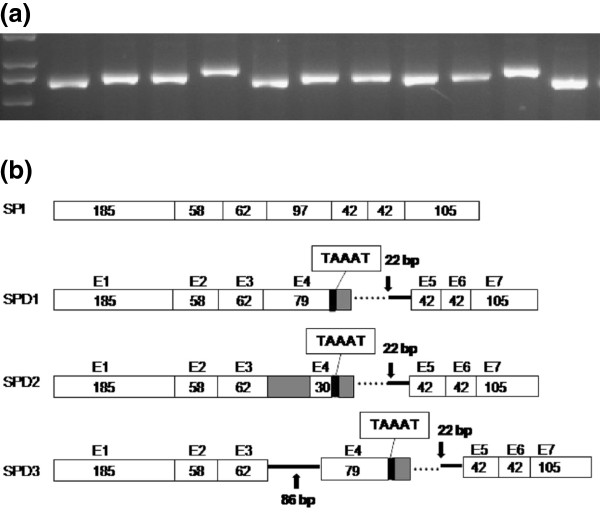
Results: We analyzed the BrFLC2 sequence in nine B. rapa accessions, and identified a 57-bp insertion/deletion (InDel) across exon 4 and intron 4 resulting in a non-functional allele. In total, three types of transcripts were identified for this mutated BrFLC2 allele. The InDel was used to develop a PCR-based marker, which was used to screen a collection of 159 B. rapa accessions. The deletion genotype was present only in oil-type B. rapa, including ssp. oleifera and ssp. tricolaris, and not in other subspecies. The deletion genotype was significantly correlated with variation in flowering time. In contrast, the reported splicing site variation in BrFLC1, which also leads to a non-functional locus, was detected but not correlated with variation in flowering time in oil-type B. rapa, although it was correlated with variation in flowering time in vegetable-type B. rapa.
Conclusions: Our results suggest that the naturally occurring deletion mutation across exon 4 and intron 4 in BrFLC2 gene contributes greatly to variation in flowering time in oil-type B. rapa. The observed different relationship between BrFLC1 or BrFLC2 and flowering time variation indicates that the control of flowering time has evolved separately between oil-type and vegetable-type B. rapa groups.
Figures
Similar articles
-
BrFLC5: a weak regulator of flowering time in Brassica rapa.Theor Appl Genet. 2018 Oct;131(10):2107-2116. doi: 10.1007/s00122-018-3139-x. Epub 2018 Jul 14. Theor Appl Genet. 2018. PMID: 30008108
-
A naturally occurring splicing site mutation in the Brassica rapa FLC1 gene is associated with variation in flowering time.J Exp Bot. 2009;60(4):1299-308. doi: 10.1093/jxb/erp010. Epub 2009 Feb 3. J Exp Bot. 2009. PMID: 19190098 Free PMC article.
-
Delayed flowering time in Arabidopsis and Brassica rapa by the overexpression of FLOWERING LOCUS C (FLC) homologs isolated from Chinese cabbage (Brassica rapa L.: ssp. pekinensis).Plant Cell Rep. 2007 Mar;26(3):327-36. doi: 10.1007/s00299-006-0243-1. Epub 2006 Oct 6. Plant Cell Rep. 2007. PMID: 17024448
-
Mapping and characterization of FLC homologs and QTL analysis of flowering time in Brassica oleracea.Theor Appl Genet. 2007 Feb;114(4):595-608. doi: 10.1007/s00122-006-0460-6. Epub 2006 Nov 29. Theor Appl Genet. 2007. PMID: 17136371
-
Genome Triplication Leads to Transcriptional Divergence of FLOWERING LOCUS C Genes During Vernalization in the Genus Brassica.Front Plant Sci. 2021 Feb 9;11:619417. doi: 10.3389/fpls.2020.619417. eCollection 2020. Front Plant Sci. 2021. PMID: 33633752 Free PMC article. Review.
Cited by
-
BrFLC5: a weak regulator of flowering time in Brassica rapa.Theor Appl Genet. 2018 Oct;131(10):2107-2116. doi: 10.1007/s00122-018-3139-x. Epub 2018 Jul 14. Theor Appl Genet. 2018. PMID: 30008108
-
Non-vernalization requirement for flowering in Brassica rapa conferred by a dominant allele of FLOWERING LOCUS T.Theor Appl Genet. 2023 May 18;136(6):132. doi: 10.1007/s00122-023-04378-y. Theor Appl Genet. 2023. PMID: 37199824
-
Investigation of Brassica and its relative genomes in the post-genomics era.Hortic Res. 2022 Aug 25;9:uhac182. doi: 10.1093/hr/uhac182. eCollection 2022. Hortic Res. 2022. PMID: 36338847 Free PMC article.
-
Non-vernalization requirement in Chinese kale caused by loss of BoFLC and low expressions of its paralogs.Theor Appl Genet. 2022 Feb;135(2):473-483. doi: 10.1007/s00122-021-03977-x. Epub 2021 Oct 29. Theor Appl Genet. 2022. PMID: 34716468 Free PMC article.
-
Association Mapping of Flowering Time QTLs and Insight into Their Contributions to Rapeseed Growth Habits.Front Plant Sci. 2016 Mar 24;7:338. doi: 10.3389/fpls.2016.00338. eCollection 2016. Front Plant Sci. 2016. PMID: 27047517 Free PMC article.
References
-
- Cao J, Schneeberger K, Ossowski S, Gunther T, Bender S, Fitz J, Koenig D, Lanz C, Stegle O, Lippert C, Wang X, Ott F, Muller J, Alonso-Blanco C, Borgwardt K, Schmid KJ, Weigel D. Whole-genome sequencing of multiple Arabidopsis thaliana populations. Nat Genet. 2011;43:956–963. doi: 10.1038/ng.911. - DOI - PubMed
-
- Koornneef M, Blankestijn-de Vries H, Hanhart C, Soppe W, Peeters T. The phenotype of some late-flowering mutants is enhanced by a locus on chromosome 5 that is not effective in the Landsberg erecta wild-type. Plant J. 1994;6:911–919. doi: 10.1046/j.1365-313X.1994.6060911.x. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
