

Late-stage maturation of the Rieske Fe/S protein: Mzm1 stabilizes Rip1 but does not facilitate its translocation by the AAA ATPase Bcs1
- PMID: 22927643
- PMCID: PMC3486142
- DOI: 10.1128/MCB.00441-12
Late-stage maturation of the Rieske Fe/S protein: Mzm1 stabilizes Rip1 but does not facilitate its translocation by the AAA ATPase Bcs1
Abstract
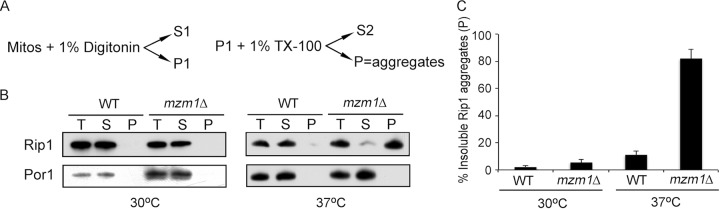
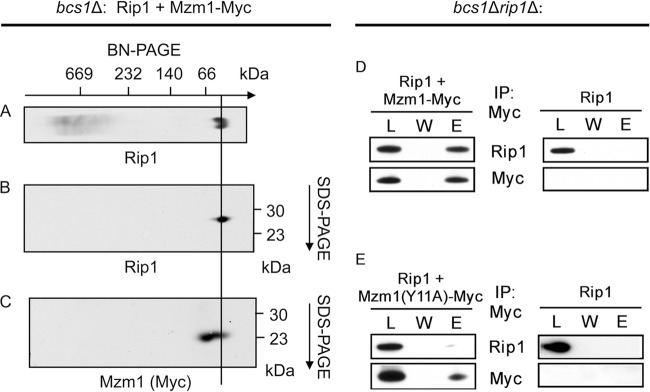
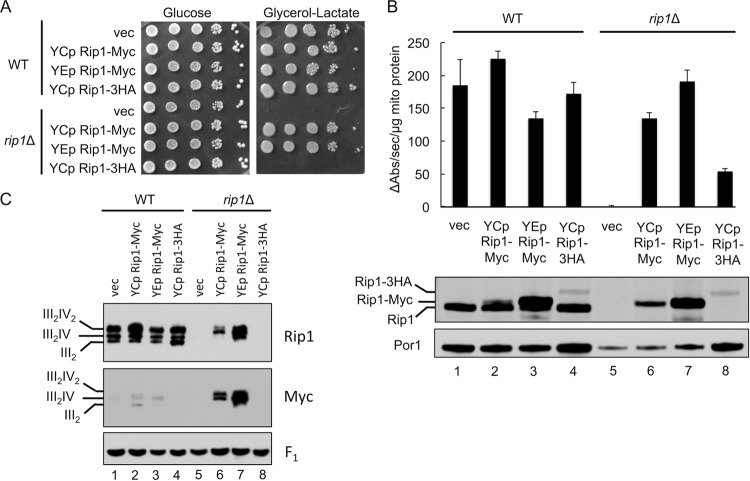
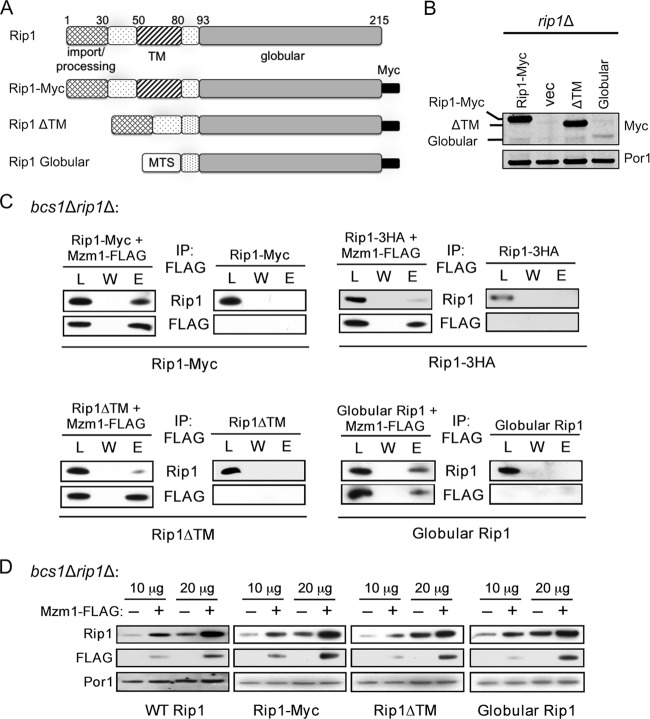
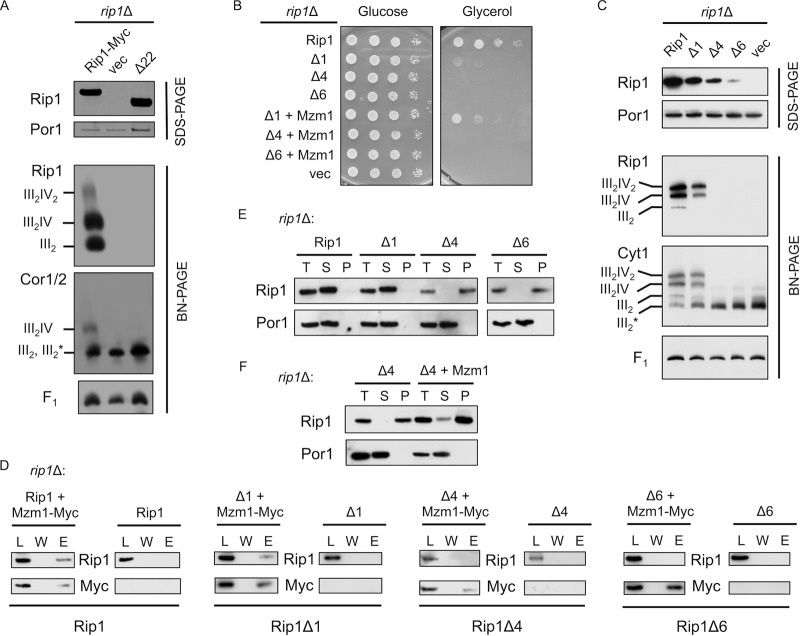
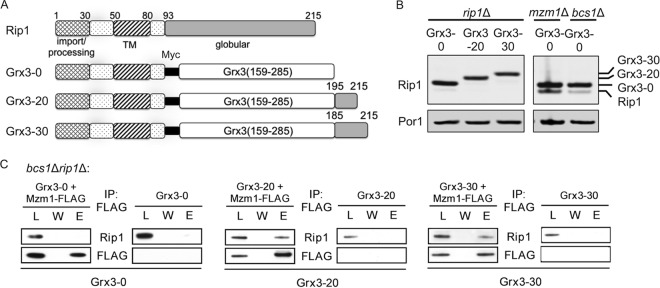
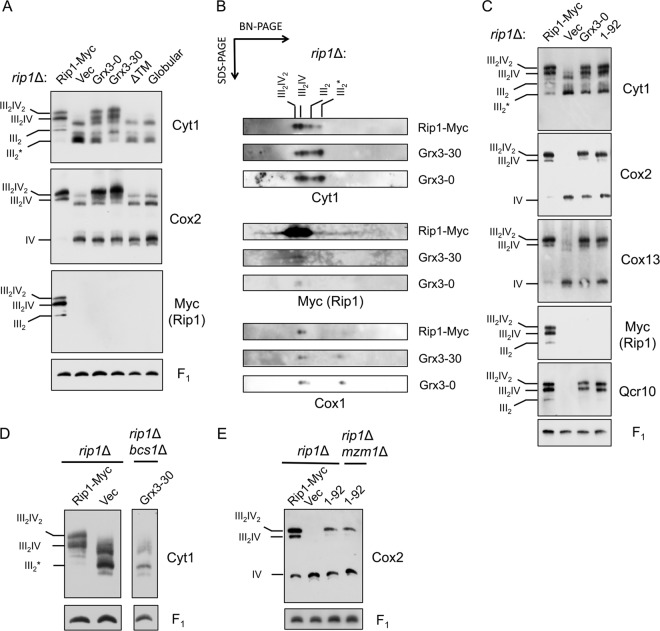
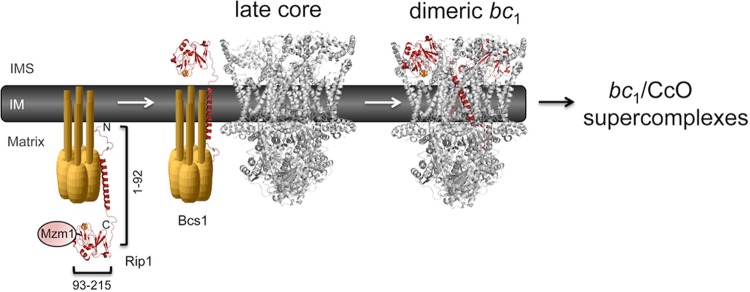
The final step in the assembly of the ubiquinol-cytochrome c reductase or bc(1) complex involves the insertion of the Rieske Fe/S cluster protein, Rip1. Maturation of Rip1 occurs within the mitochondrial matrix prior to its translocation across the inner membrane (IM) in a process mediated by the Bcs1 ATPase and subsequent insertion into the bc(1) complex. Here we show that the matrix protein Mzm1 functions as a Rip1 chaperone, stabilizing Rip1 prior to the translocation step. In the absence of Mzm1, Rip1 is prone to either proteolytic degradation or temperature-induced aggregation. A series of Rip1 truncations were engineered to probe motifs necessary for Mzm1 interaction and Bcs1-mediated translocation of Rip1. The Mzm1 interaction with Rip1 persists in Rip1 variants lacking its transmembrane domain or containing only its C-terminal globular Fe/S domain. Replacement of the globular domain of Rip1 with that of the heterologous folded protein Grx3 abrogated Mzm1 interaction; however, appending the C-terminal 30 residues of Rip1 to the Rip1-Grx3 chimera restored Mzm1 interaction. The Rip1-Grx3 chimera and a Rip1 truncation containing only the N-terminal 92 residues each induced stabilization of the bc(1):cytochrome oxidase supercomplex in a Bcs1-dependent manner. However, the Rip1 variants were not stably associated with the supercomplex. The induced supercomplex stabilization by the Rip1 N terminus was independent of Mzm1.
Figures
References
-
- Bachmann J, Bauer B, Zwicker K, Ludwig B, Anderka O. 2006. The Rieske protein from Paracoccus denitrificans is inserted into the cytoplasmic membrane by the twin-arginine translocase. FEBS J. 273: 4817–4830 - PubMed
-
- Brandt U, Uribe S, Schagger H, Trumpower BL. 1994. Isolation and characterization of QCR10, the nuclear gene encoding the 8.5-kDa subunit 10 of the Saccharomyces cerevisiae cytochrome bc1 complex. J. Biol. Chem. 269: 12947–12953 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous