

A novel virulence strategy for Pseudomonas aeruginosa mediated by an autotransporter with arginine-specific aminopeptidase activity
- PMID: 22927813
- PMCID: PMC3426542
- DOI: 10.1371/journal.ppat.1002854
A novel virulence strategy for Pseudomonas aeruginosa mediated by an autotransporter with arginine-specific aminopeptidase activity
Abstract
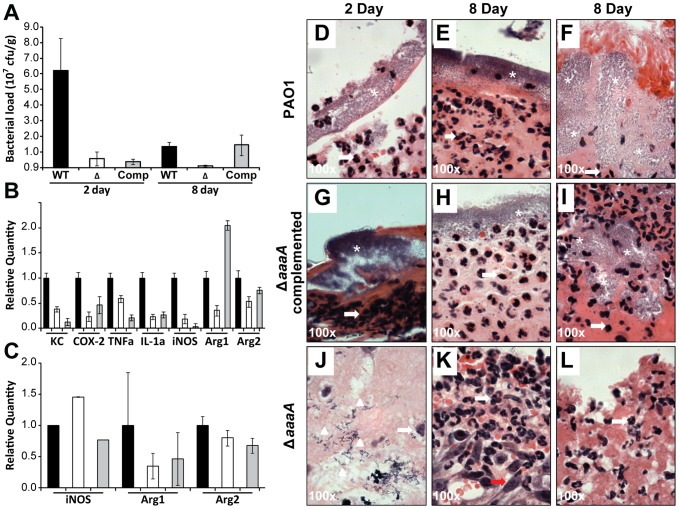
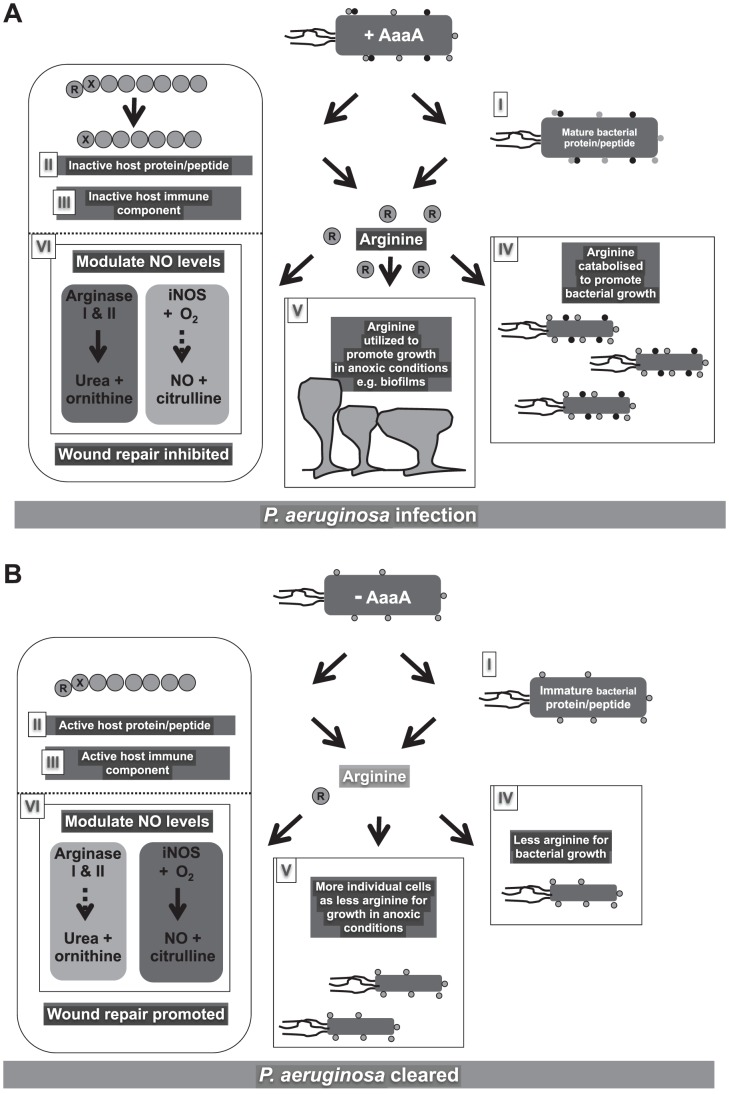
The opportunistic human pathogen, Pseudomonas aeruginosa, is a major cause of infections in chronic wounds, burns and the lungs of cystic fibrosis patients. The P. aeruginosa genome encodes at least three proteins exhibiting the characteristic three domain structure of autotransporters, but much remains to be understood about the functions of these three proteins and their role in pathogenicity. Autotransporters are the largest family of secreted proteins in Gram-negative bacteria, and those characterised are virulence factors. Here, we demonstrate that the PA0328 autotransporter is a cell-surface tethered, arginine-specific aminopeptidase, and have defined its active site by site directed mutagenesis. Hence, we have assigned PA0328 with the name AaaA, for arginine-specific autotransporter of P. aeruginosa. We show that AaaA provides a fitness advantage in environments where the sole source of nitrogen is peptides with an aminoterminal arginine, and that this could be important for establishing an infection, as the lack of AaaA led to attenuation in a mouse chronic wound infection which correlated with lower levels of the cytokines TNFα, IL-1α, KC and COX-2. Consequently AaaA is an important virulence factor playing a significant role in the successful establishment of P. aeruginosa infections.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
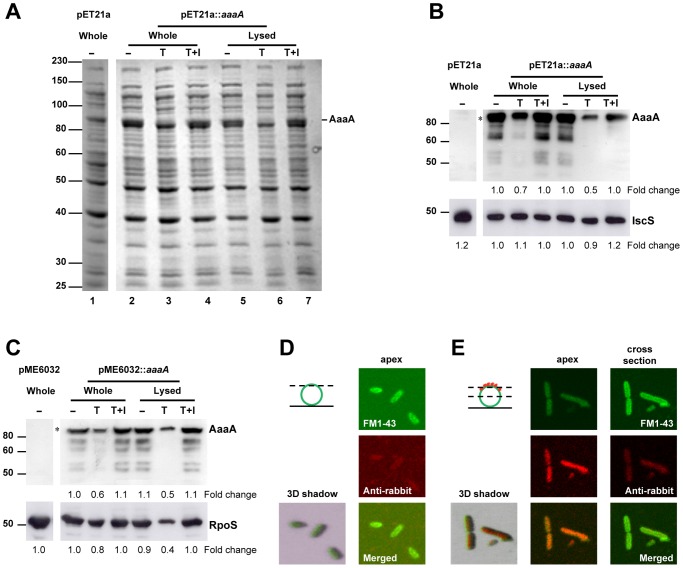
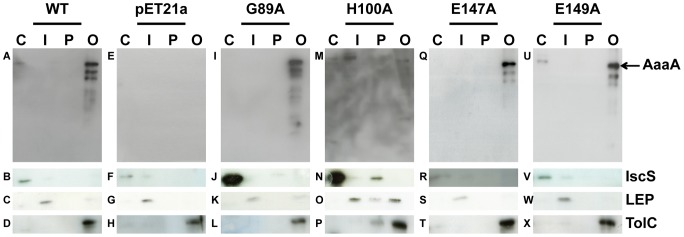
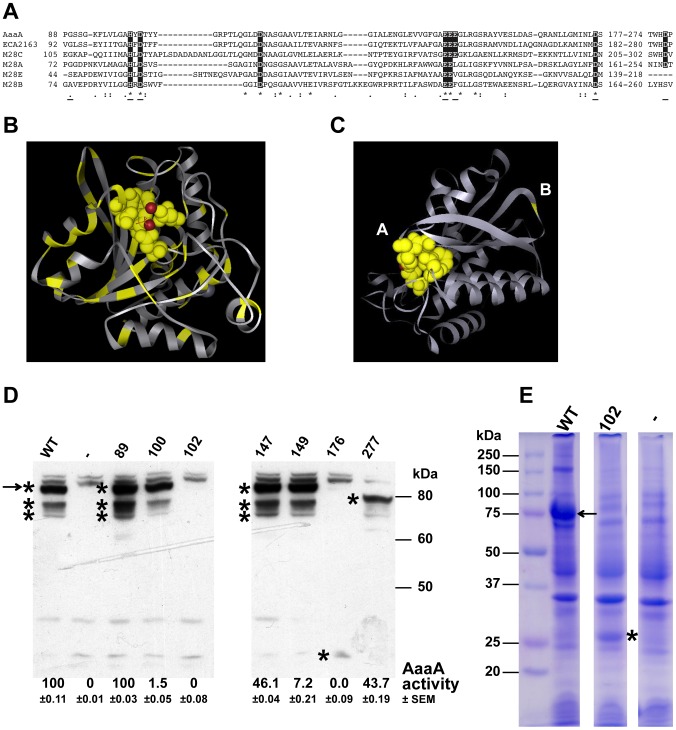


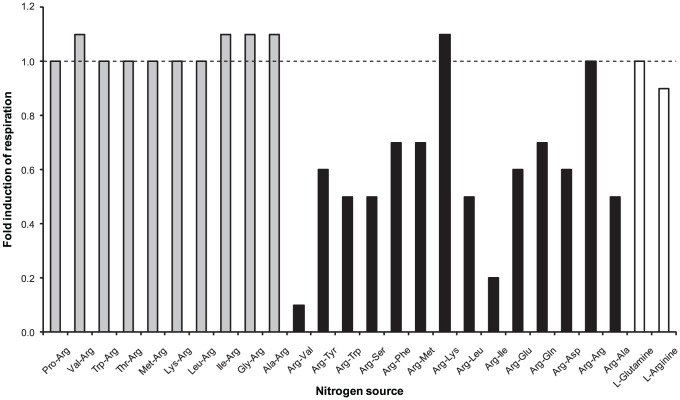
References
-
- Kerr KG, Snelling AM (2009) Pseudomonas aeuginosa: a formidable and ever-present adversary. J Hosp Infect 73: 338–344. - PubMed
-
- George AM, Jones PM, Middleton PG (2009) Cystic fibrosis infections: treatment strategies and prospects. FEMS Microbiol Lett 300: 153–164. - PubMed
-
- Doring G, Conway S, Heijerman H, Hodson M, Hoiby N, et al. (2000) Antibiotic therapy against Pseudomonas aeruginosa in cystic fibrosis: a European consensus. Eur Respir J 16: 749–467. - PubMed
-
- El Solh AA, Alhajhusain A (2009) Update on the treatment of Pseudomonas aeruginosa pneumonia. J Antimicrob Chemother 64: 229–238. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
