

Energy demand of synaptic transmission at the hippocampal Schaffer-collateral synapse
- PMID: 22929439
- PMCID: PMC3493999
- DOI: 10.1038/jcbfm.2012.116
Energy demand of synaptic transmission at the hippocampal Schaffer-collateral synapse
Abstract
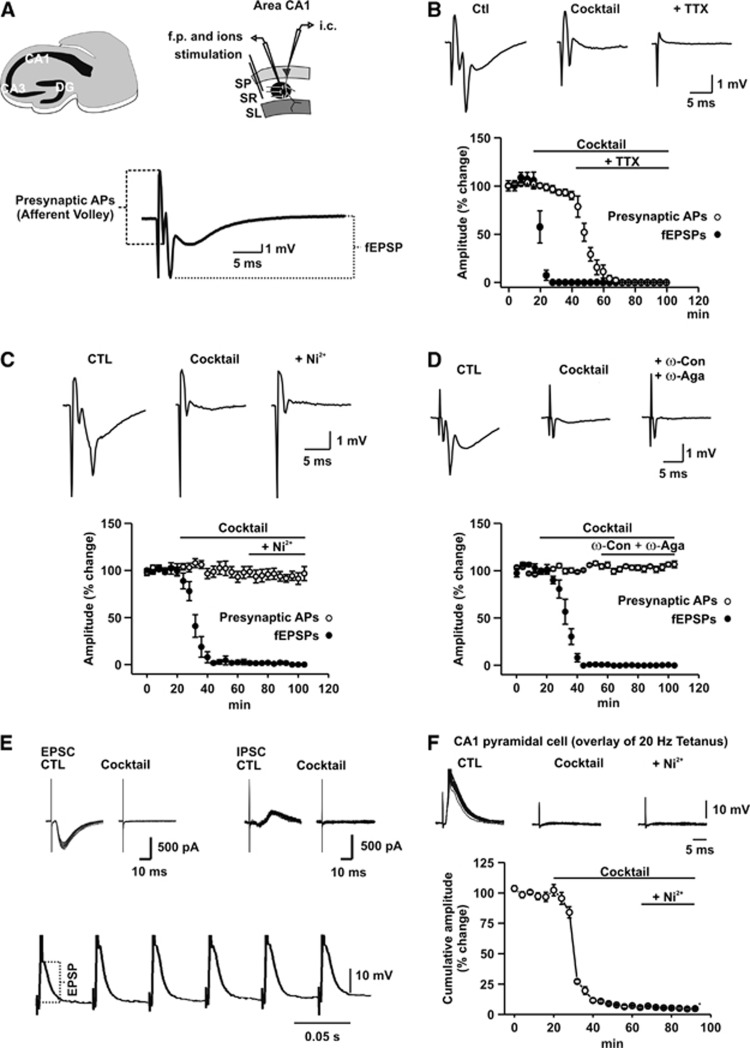
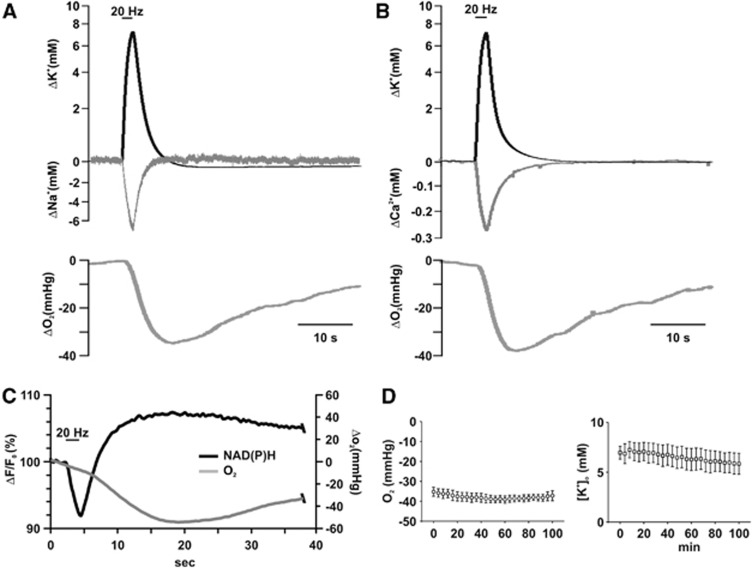
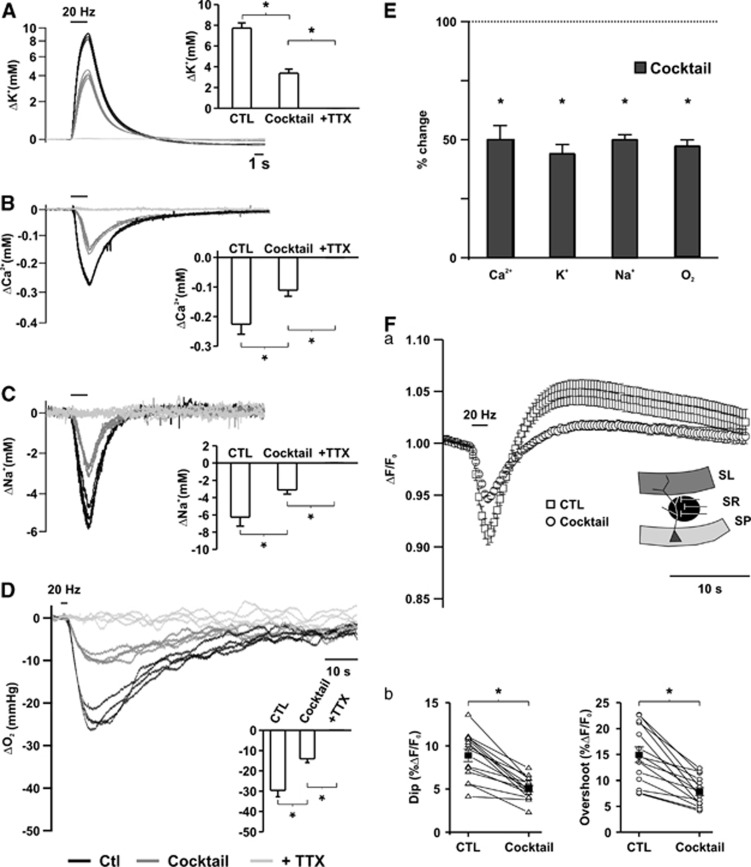
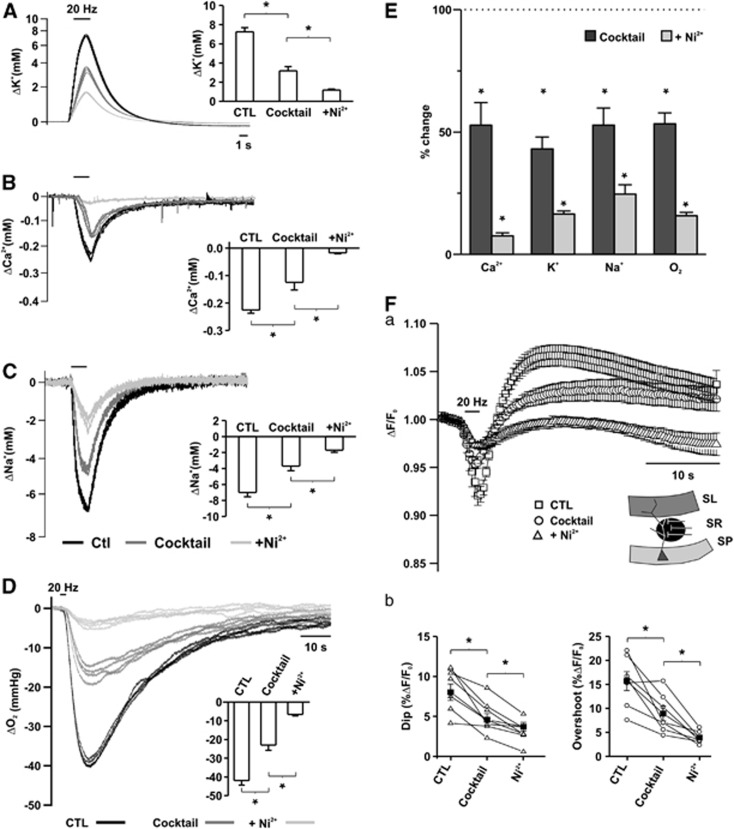
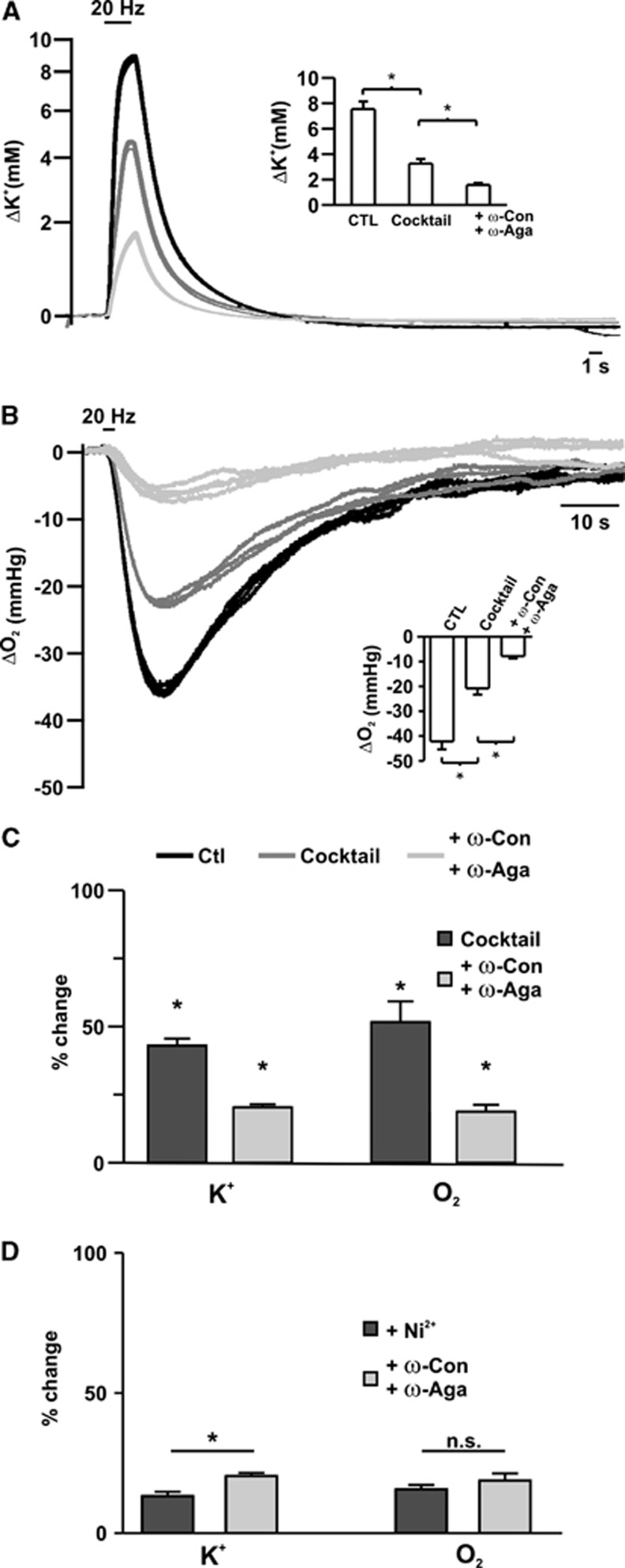
Neuroenergetic models of synaptic transmission predicted that energy demand is highest for action potentials (APs) and postsynaptic ion fluxes, whereas the presynaptic contribution is rather small. Here, we addressed the question of energy consumption at Schaffer-collateral synapses. We monitored stimulus-induced changes in extracellular potassium, sodium, and calcium concentration while recording partial oxygen pressure (pO(2)) and NAD(P)H fluorescence. Blockade of postsynaptic receptors reduced ion fluxes as well as pO(2) and NAD(P)H transients by ∼50%. Additional blockade of transmitter release further reduced Na(+), K(+), and pO(2) transients by ∼30% without altering presynaptic APs, indicating considerable contribution of Ca(2+)-removal, transmitter and vesicle turnover to energy consumption.
Figures
References
-
- Attwell D, Gibb A. Neuroenergetics and the kinetic design of excitatory synapses. Nat Rev Neurosci. 2005;6:841–849. - PubMed
-
- Attwell D, Laughlin SB. An energy budget for signaling in the grey matter of the brain. J Cereb Blood Flow Metab. 2001;21:1133–1145. - PubMed
-
- Alle H, Roth A, Geiger JR. Energy-efficient action potentials in hippocampal mossy fibers. Science. 2009;325:1405–1408. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
