

miRNA-21 is developmentally regulated in mouse brain and is co-expressed with SOX2 in glioma
- PMID: 22931209
- PMCID: PMC3517377
- DOI: 10.1186/1471-2407-12-378
miRNA-21 is developmentally regulated in mouse brain and is co-expressed with SOX2 in glioma
Abstract
Background: MicroRNAs (miRNAs) and their role during tumor development have been studied in great detail during the last decade, albeit their expression pattern and regulation during normal development are however not so well established. Previous studies have shown that miRNAs are differentially expressed in solid human tumors. Platelet-derived growth factor (PDGF) signaling is known to be involved in normal development of the brain as well as in malignant primary brain tumors, gliomas, but the complete mechanism is still lacking. We decided to investigate the expression of the oncogenic miR-21 during normal mouse development and glioma, focusing on PDGF signaling as a potential regulator of miR-21.
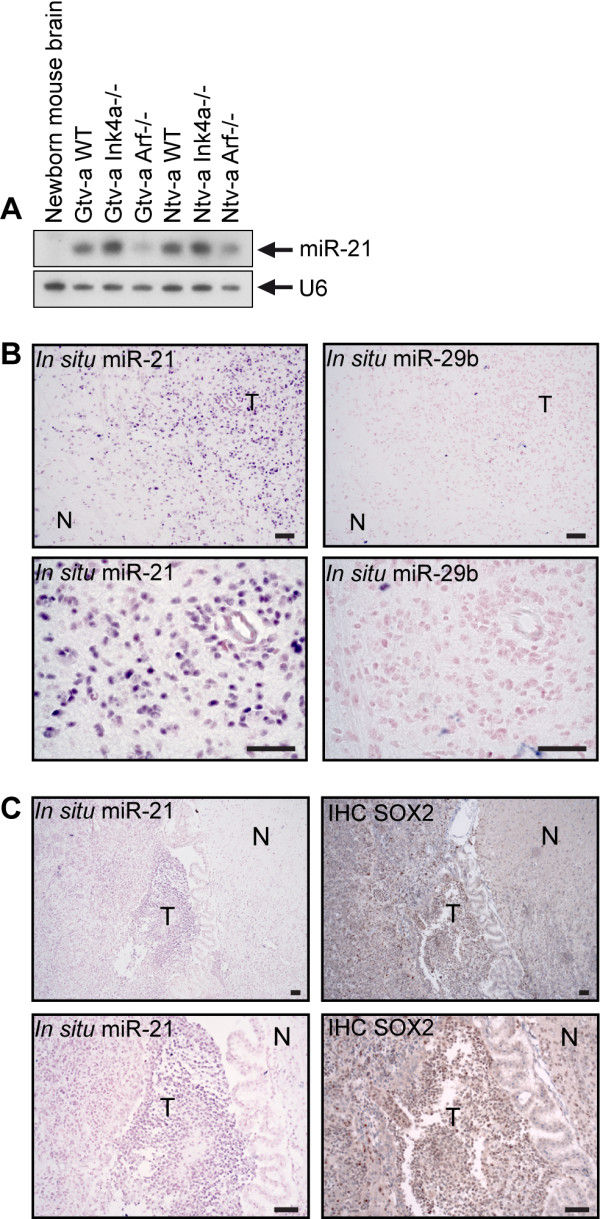
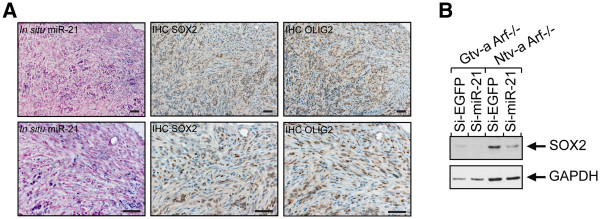
Methods: We generated mouse glioma using the RCAS/tv-a system for driving PDGF-BB expression in a cell-specific manner. Expression of miR-21 in mouse cell cultures and mouse brain were assessed using Northern blot analysis and in situ hybridization. Immunohistochemistry and Western blot analysis were used to investigate SOX2 expression. LNA-modified siRNA was used for irreversible depletion of miR-21. For inhibition of PDGF signaling Gleevec (imatinib mesylate), Rapamycin and U0126, as well as siRNA were used. Statistical significance was calculated using double-sided unpaired Student's t-test.
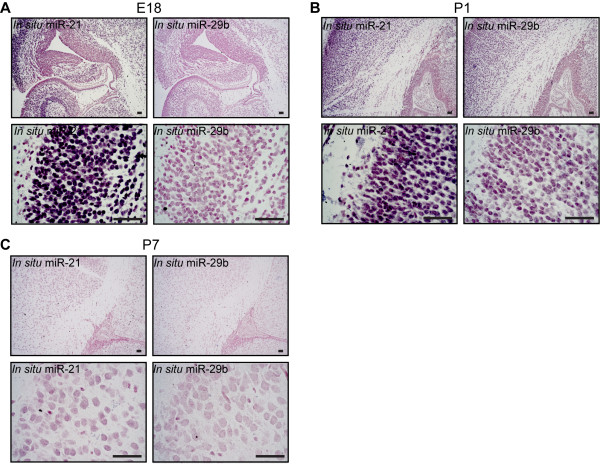
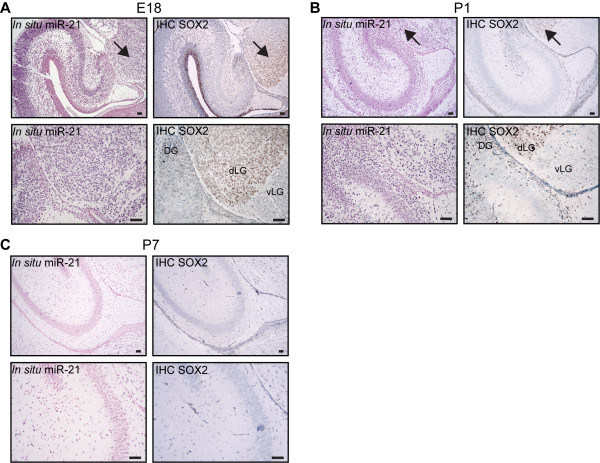
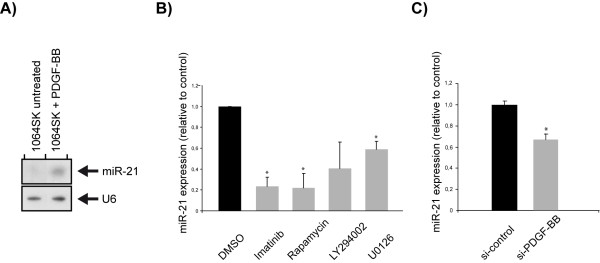
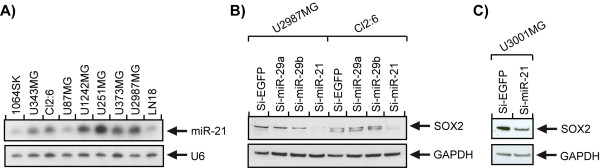
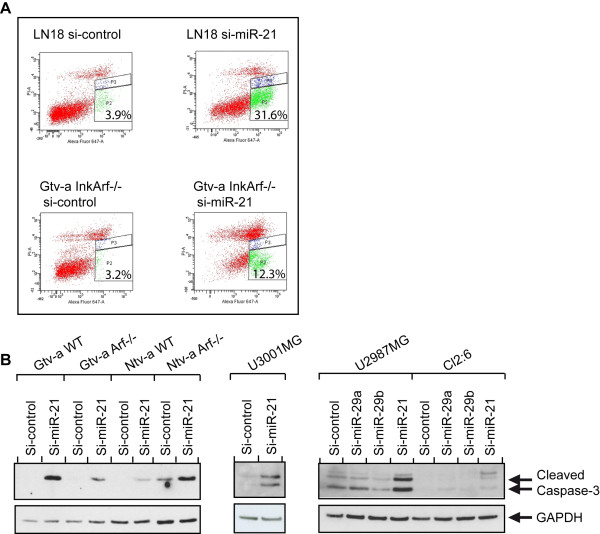
Results: We identified miR-21 to be highly expressed during embryonic and newborn brain development followed by a gradual decrease until undetectable at postnatal day 7 (P7), this pattern correlated with SOX2 expression. Furthermore, miR-21 and SOX2 showed up-regulation and overlapping expression pattern in RCAS/tv-a generated mouse brain tumor specimens. Upon irreversible depletion of miR-21 the expression of SOX2 was strongly diminished in both mouse primary glioma cultures and human glioma cell lines. Interestingly, in normal fibroblasts the expression of miR-21 was induced by PDGF-BB, and inhibition of PDGF signaling in mouse glioma primary cultures resulted in suppression of miR-21 suggesting that miR-21 is indeed regulated by PDGF signaling.
Conclusions: Our data show that miR-21 and SOX2 are tightly regulated already during embryogenesis and define a distinct population with putative tumor cell of origin characteristics. Furthermore, we believe that miR-21 is a mediator of PDGF-driven brain tumors, which suggests miR-21 as a promising target for treatment of glioma.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
