

Comparative genomics guided discovery of two missing archaeal enzyme families involved in the biosynthesis of the pterin moiety of tetrahydromethanopterin and tetrahydrofolate
- PMID: 22931285
- PMCID: PMC3500442
- DOI: 10.1021/cb300342u
Comparative genomics guided discovery of two missing archaeal enzyme families involved in the biosynthesis of the pterin moiety of tetrahydromethanopterin and tetrahydrofolate
Abstract
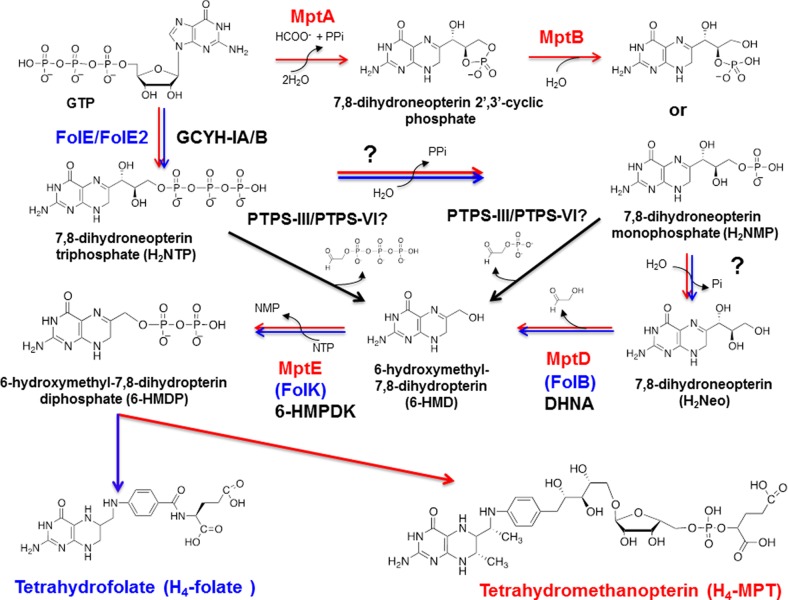
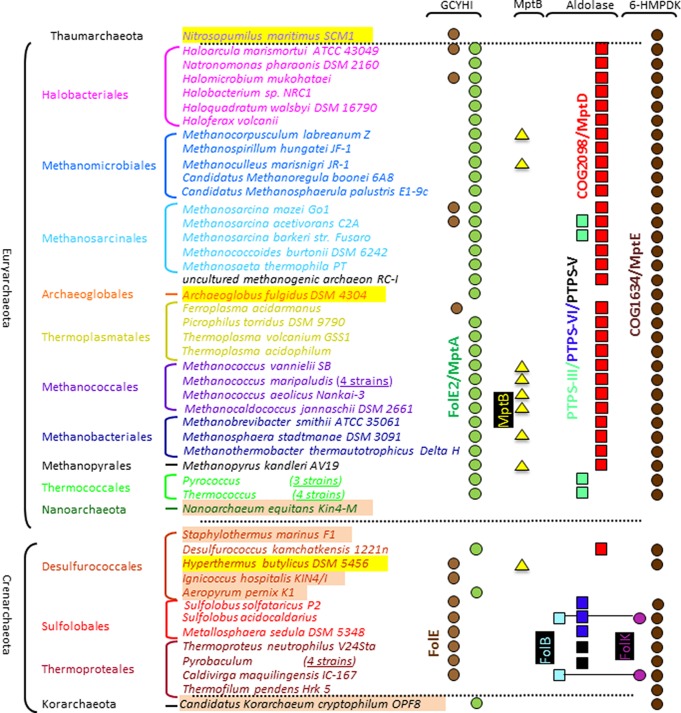
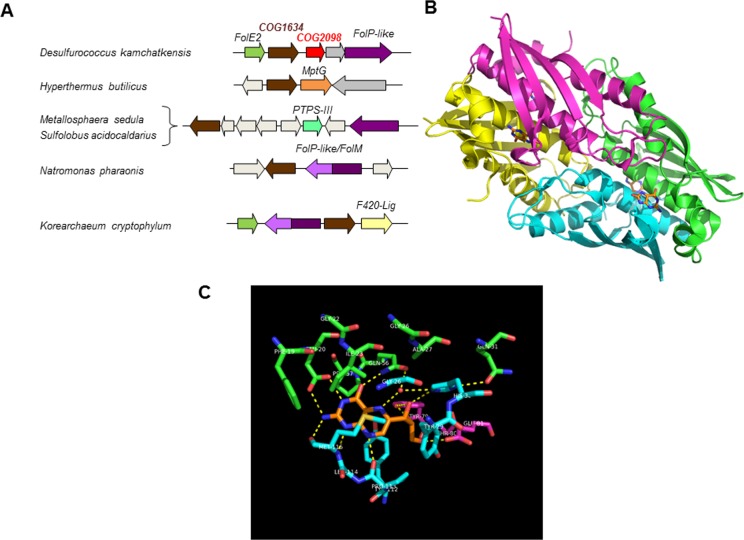
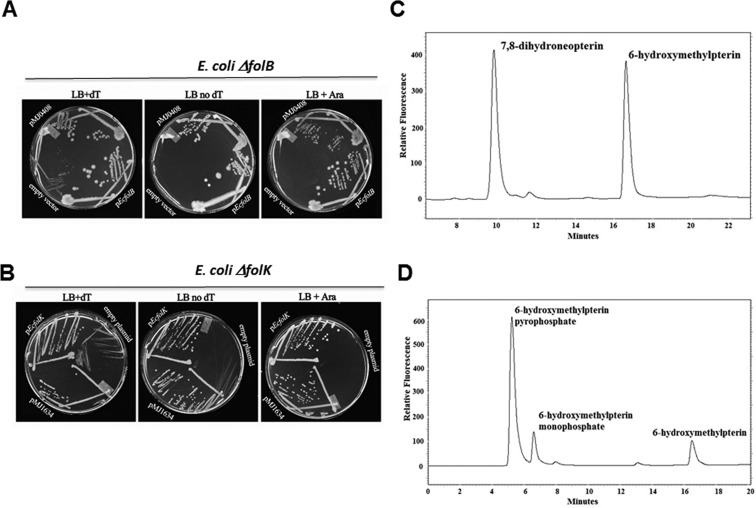
C-1 carriers are essential cofactors in all domains of life, and in Archaea, these can be derivatives of tetrahydromethanopterin (H(4)-MPT) or tetrahydrofolate (H(4)-folate). Their synthesis requires 6-hydroxymethyl-7,8-dihydropterin diphosphate (6-HMDP) as the precursor, but the nature of pathways that lead to its formation were unknown until the recent discovery of the GTP cyclohydrolase IB/MptA family that catalyzes the first step, the conversion of GTP to dihydroneopterin 2',3'-cyclic phosphate or 7,8-dihydroneopterin triphosphate [El Yacoubi, B.; et al. (2006) J. Biol. Chem., 281, 37586-37593 and Grochowski, L. L.; et al. (2007) Biochemistry46, 6658-6667]. Using a combination of comparative genomics analyses, heterologous complementation tests, and in vitro assays, we show that the archaeal protein families COG2098 and COG1634 specify two of the missing 6-HMDP synthesis enzymes. Members of the COG2098 family catalyze the formation of 6-hydroxymethyl-7,8-dihydropterin from 7,8-dihydroneopterin, while members of the COG1634 family catalyze the formation of 6-HMDP from 6-hydroxymethyl-7,8-dihydropterin. The discovery of these missing genes solves a long-standing mystery and provides novel examples of convergent evolutions where proteins of dissimilar architectures perform the same biochemical function.
Figures
References
-
- Overbeek R.; Begley T.; Butler R. M.; Choudhuri J. V.; Chuang H. Y.; Cohoon M.; de Crécy-Lagard V.; Diaz N.; Disz T.; Edwards R.; Fonstein M.; Frank E. D.; Gerdes S.; Glass E. M.; Goesmann A.; Hanson A.; Iwata-Reuyl D.; Jensen R.; Jamshidi N.; Krause L.; Kubal M.; Larsen N.; Linke B.; McHardy A. C.; Meyer F.; Neuweger H.; Olsen G.; Olson R.; Osterman A.; Portnoy V.; Pusch G. D.; Rodionov D. A.; Ruckert C.; Steiner J.; Stevens R.; Thiele I.; Vassieva O.; Ye Y.; Zagnitko O.; Vonstein V. (2005) The subsystems approach to genome annotation and its use in the project to annotate 1000 genomes. Nucleic Acids Res. 33, 5691–5702. - PMC - PubMed
-
- Henry C. S.; Overbeek R.; Xia F.; Best A. A.; Glass E.; Gilbert J.; Larsen P.; Edwards R.; Disz T.; Meyer F.; Vonstein V.; DeJongh M.; Bartels D.; Desai N.; D’Souza M.; Devoid S.; Keegan K. P.; Olson R.; Wilke A.; Wilkening J.; Stevens R. L. (2011) Connecting genotype to phenotype in the era of high-throughput sequencing. Biochim. Biophys. Acta 1810, 967–977. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
