

Small signaling peptides in Arabidopsis development: how cells communicate over a short distance
- PMID: 22932676
- PMCID: PMC3462626
- DOI: 10.1105/tpc.112.099010
Small signaling peptides in Arabidopsis development: how cells communicate over a short distance
Abstract
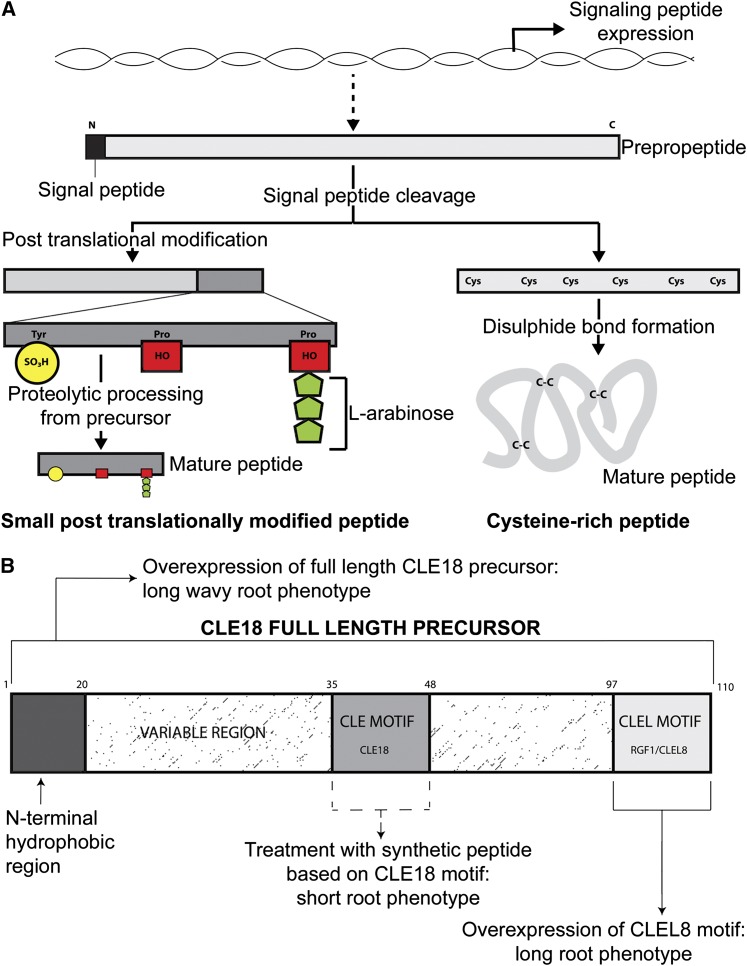
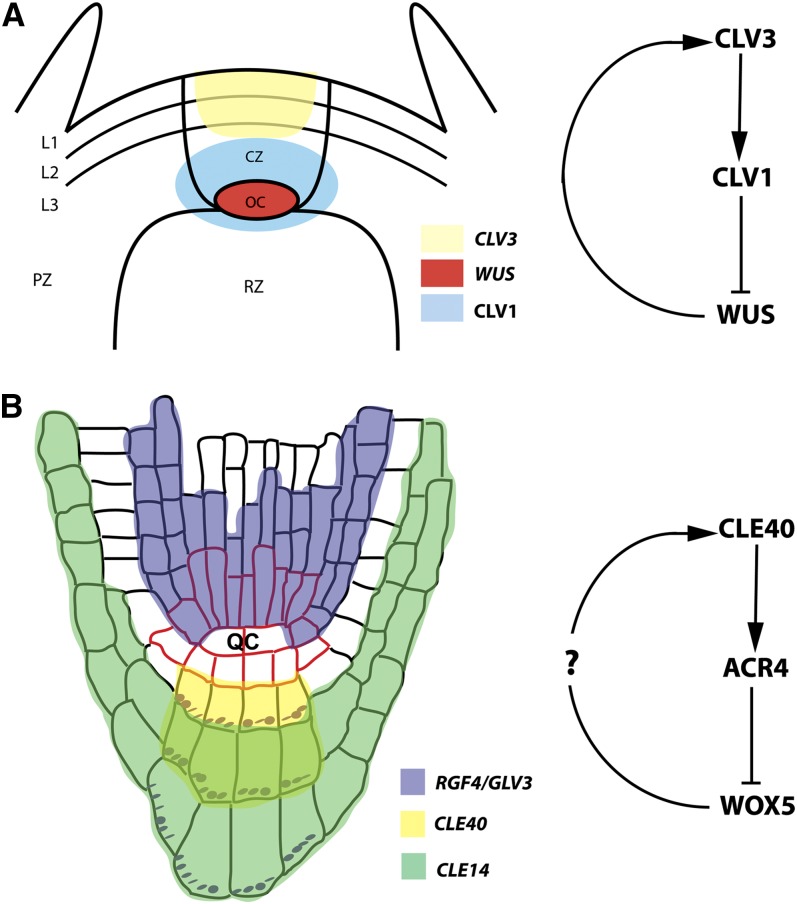
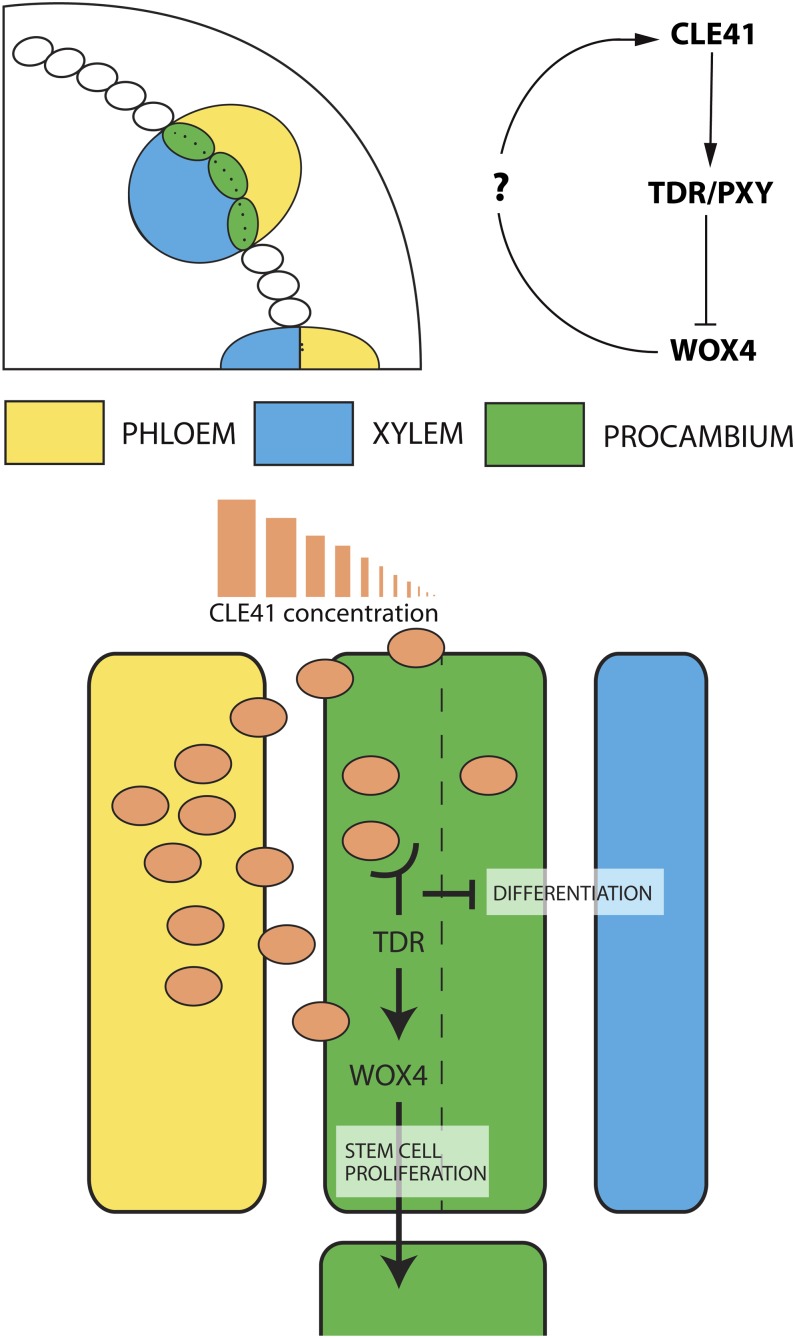
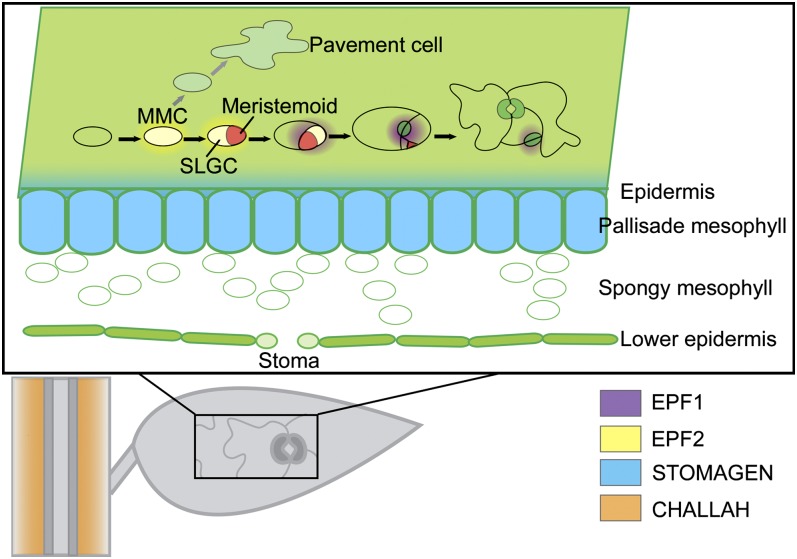
To sustain plants' postembryonic growth and development in a structure of cells fixed in cell walls, a tightly controlled short distance cell-cell communication is required. The focus on phytohormones, such as auxin, has historically overshadowed the importance of small peptide signals, but it is becoming clear that secreted peptide signals are important in cell-cell communication to coordinate and integrate cellular functions. However, of the more than 1000 potential secreted peptides, so far only very few have been functionally characterized or matched to a receptor. Here, we will describe our current knowledge on how small peptide signals can be identified, how they are modified and processed, which roles they play in Arabidopsis thaliana development, and through which receptors they act.
Figures
References
-
- Abrash E.B., Bergmann D.C. (2010). Regional specification of stomatal production by the putative ligand CHALLAH. Development 137: 447–455 - PubMed
-
- Aida M., Beis D., Heidstra R., Willemsen V., Blilou I., Galinha C., Nussaume L., Noh Y.S., Amasino R., Scheres B. (2004). The PLETHORA genes mediate patterning of the Arabidopsis root stem cell niche. Cell 119: 109–120 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
