

SSTR2 is the functionally dominant somatostatin receptor in human pancreatic β- and α-cells
- PMID: 22932785
- PMCID: PMC3492856
- DOI: 10.1152/ajpendo.00207.2012
SSTR2 is the functionally dominant somatostatin receptor in human pancreatic β- and α-cells
Abstract
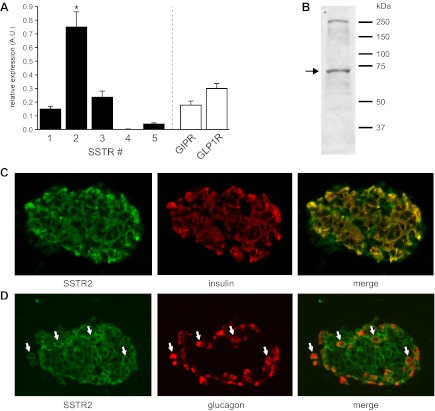
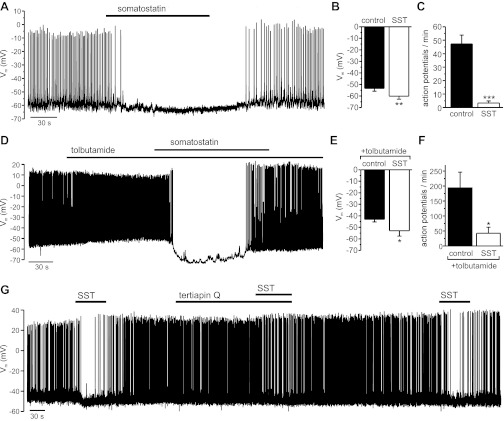
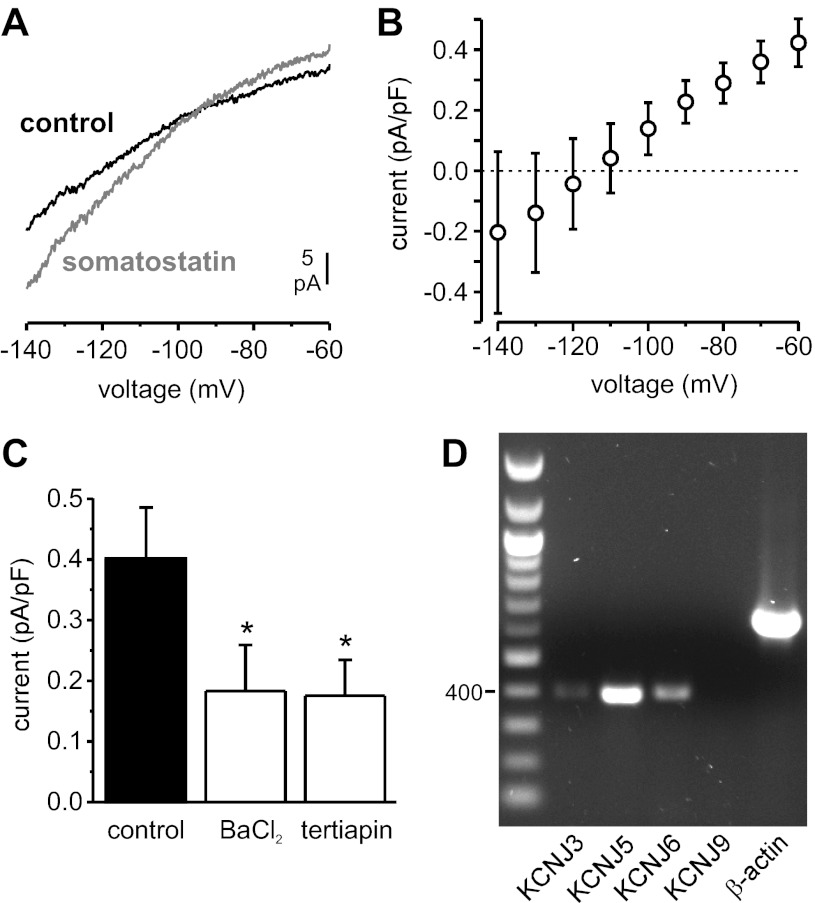
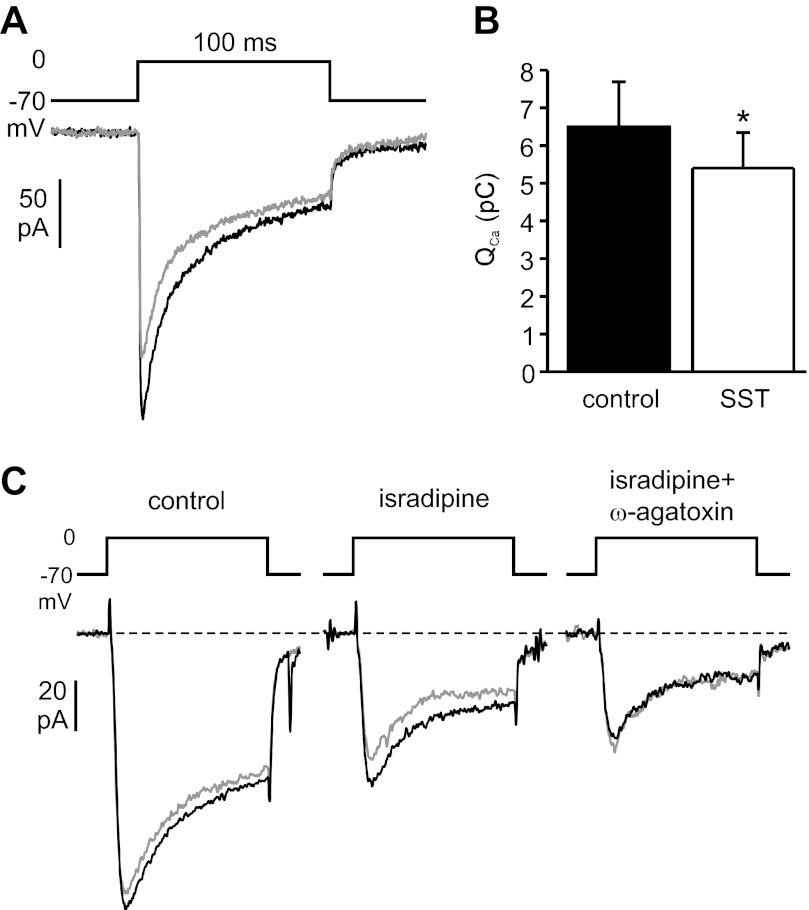
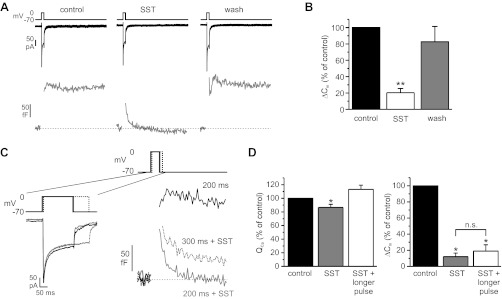
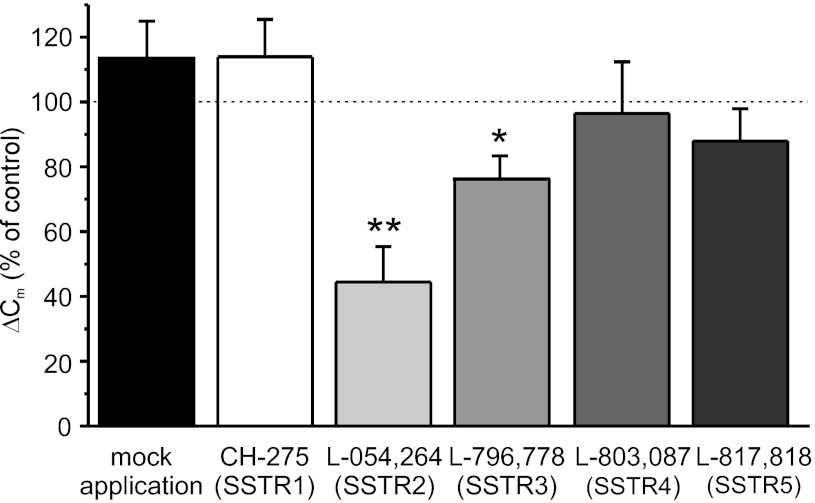
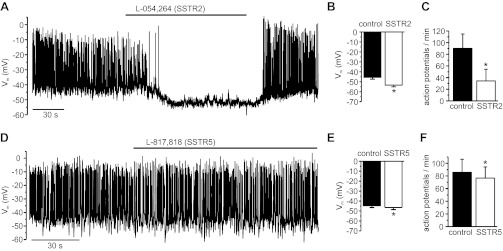
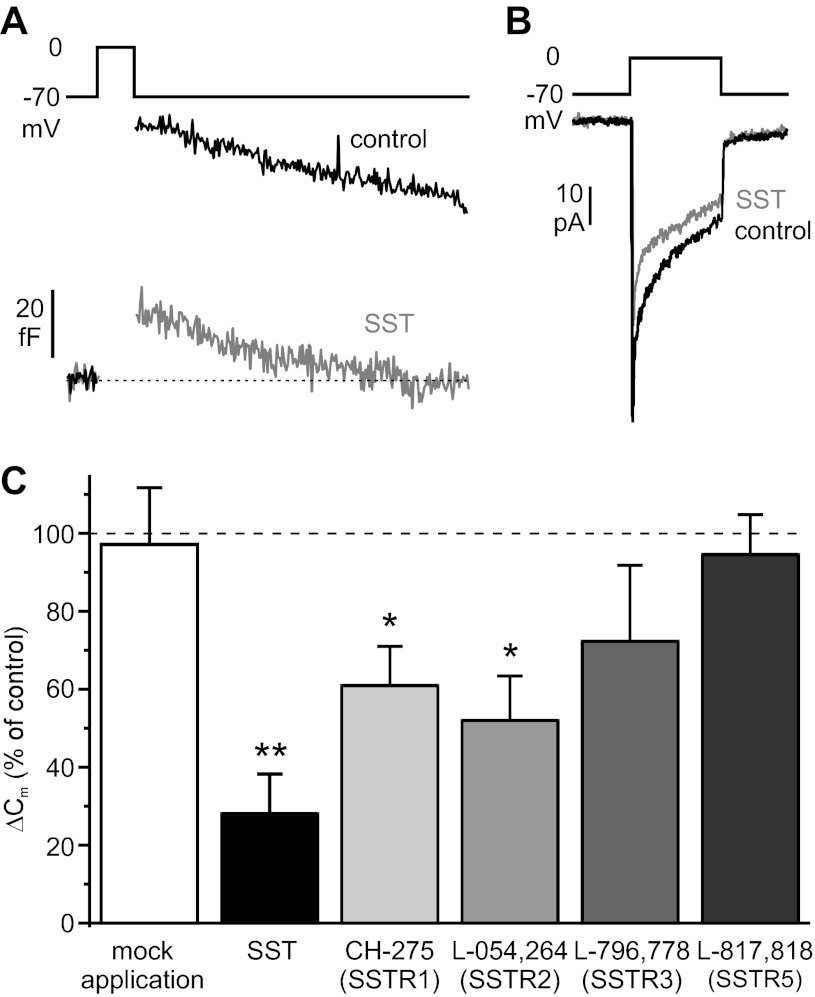
Somatostatin-14 (SST) inhibits insulin and glucagon secretion by activating G protein-coupled somatostatin receptors (SSTRs), of which five isoforms exist (SSTR1-5). In mice, the effects on pancreatic β-cells are mediated by SSTR5, whereas α-cells express SSTR2. In both cell types, SSTR activation results in membrane hyperpolarization and suppression of exocytosis. Here, we examined the mechanisms by which SST inhibits secretion from human β- and α-cells and the SSTR isoforms mediating these effects. Quantitative PCR revealed high expression of SSTR2, with lower levels of SSTR1, SSTR3, and SSTR5, in human islets. Immunohistochemistry showed expression of SSTR2 in both β- and α-cells. SST application hyperpolarized human β-cells and inhibited action potential firing. The membrane hyperpolarization was unaffected by tolbutamide but antagonized by tertiapin-Q, a blocker of G protein-gated inwardly rectifying K⁺ channels (GIRK). The effect of SST was mimicked by an SSTR2-selective agonist, whereas a SSTR5 agonist was marginally effective. SST strongly (>70%) reduced depolarization-evoked exocytosis in both β- and α-cells. A slightly weaker inhibition was observed in both cell types after SSTR2 activation. SSTR3- and SSTR1-selective agonists moderately reduced the exocytotic responses in β- and α-cells, respectively, whereas SSTR4- and SSTR5-specific agonists were ineffective. SST also reduced voltage-gated P/Q-type Ca²⁺ currents in β-cells, but normalization of Ca²⁺ influx to control levels by prolonged depolarizations only partially restored exocytosis. We conclude that SST inhibits secretion from both human β- and α-cells by activating GIRK and suppressing electrical activity, reducing P/Q-type Ca²⁺ currents, and directly inhibiting exocytosis. These effects are mediated predominantly by SSTR2 in both cell types.
Figures
References
-
- Barnett P. Somatostatin and somatostatin receptor physiology. Endocrine 20: 255–264, 2003 - PubMed
-
- Braun M, Ramracheya R, Amisten S, Bengtsson M, Moritoh Y, Zhang Q, Johnson PR, Rorsman P. Somatostatin release, electrical activity, membrane currents and exocytosis in human pancreatic delta cells. Diabetologia 52: 1566–1578, 2009 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous