

Metabolic hypothesis for human altriciality
- PMID: 22932870
- PMCID: PMC3458333
- DOI: 10.1073/pnas.1205282109
Metabolic hypothesis for human altriciality
Abstract
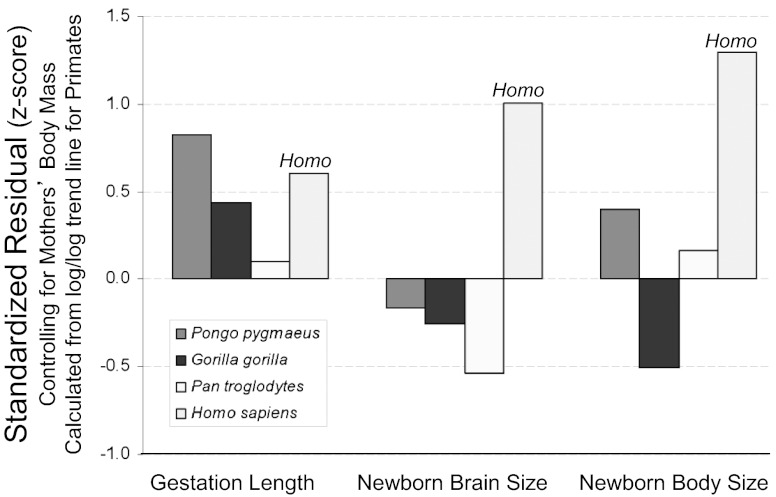
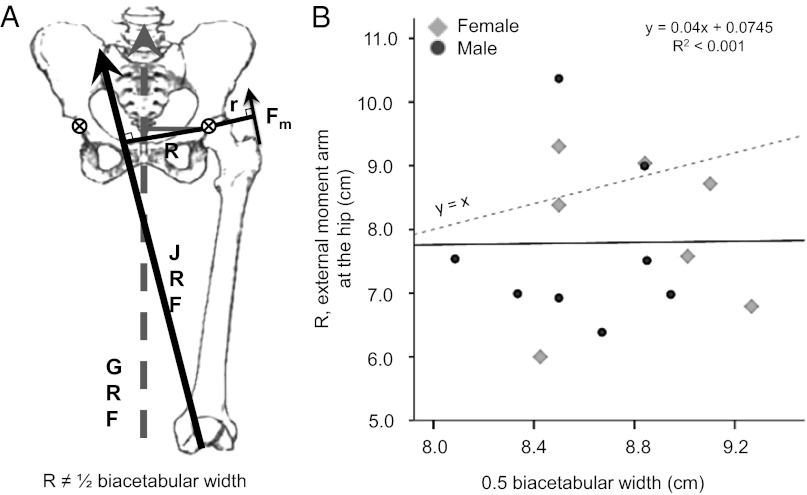
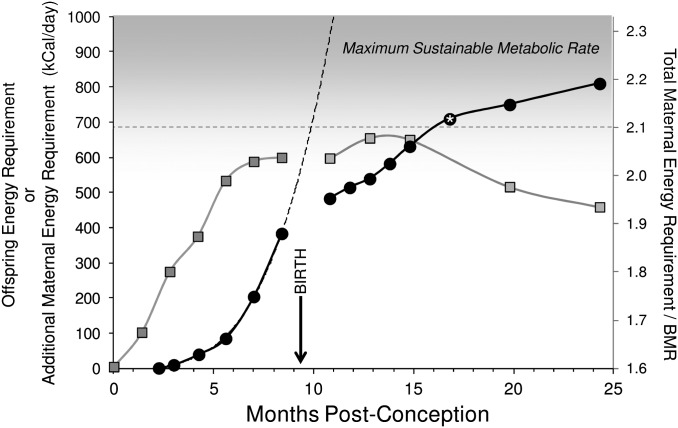
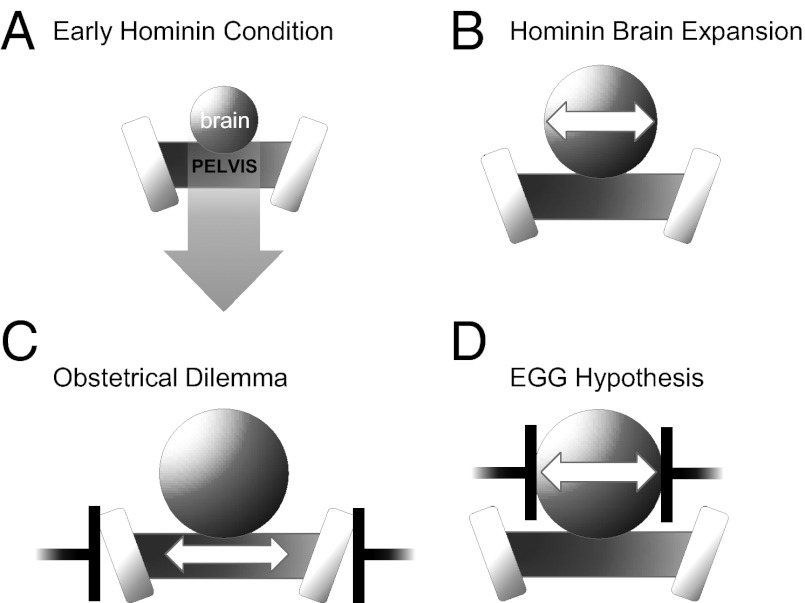
The classic anthropological hypothesis known as the "obstetrical dilemma" is a well-known explanation for human altriciality, a condition that has significant implications for human social and behavioral evolution. The hypothesis holds that antagonistic selection for a large neonatal brain and a narrow, bipedal-adapted birth canal poses a problem for childbirth; the hominin "solution" is to truncate gestation, resulting in an altricial neonate. This explanation for human altriciality based on pelvic constraints persists despite data linking human life history to that of other species. Here, we present evidence that challenges the importance of pelvic morphology and mechanics in the evolution of human gestation and altriciality. Instead, our analyses suggest that limits to maternal metabolism are the primary constraints on human gestation length and fetal growth. Although pelvic remodeling and encephalization during hominin evolution contributed to the present parturitional difficulty, there is little evidence that pelvic constraints have altered the timing of birth.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Stearns SC. The influence of size and phylogeny on patterns of covariation among life-history traits in the mammals. Oikos. 1983;41:173–187.
-
- Harvey PH, Clutton-Brock TH. Life history variation in primates. Evolution. 1985;39:559–581. - PubMed
-
- Read AE, Harvey PH. Life history differences among the eutherian radiations. J Zool (Lond) 1989;219:329–353.
-
- DeSilva J, Lesnik J. Chimpanzee neonatal brain size: Implications for brain growth in Homo erectus. J Hum Evol. 2006;51:207–212. - PubMed
-
- Portmann A. 1969. Biologische Fragmente zu einer Lehre vom Menschen [A Zoologist Looks at Humankind] (Schwabe, Basel, Germany); trans Schaefer J (1990) (Columbia University Press, New York). German.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
