

Increased hepatic glucose production in fetal sheep with intrauterine growth restriction is not suppressed by insulin
- PMID: 22933111
- PMCID: PMC3526037
- DOI: 10.2337/db11-1727
Increased hepatic glucose production in fetal sheep with intrauterine growth restriction is not suppressed by insulin
Abstract
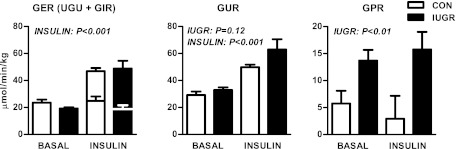
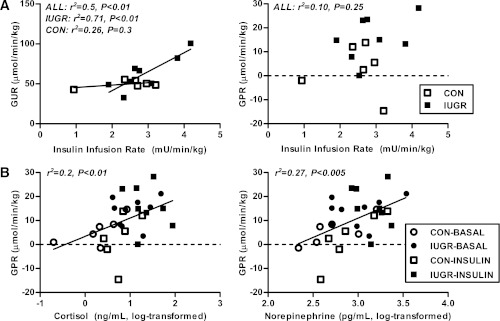
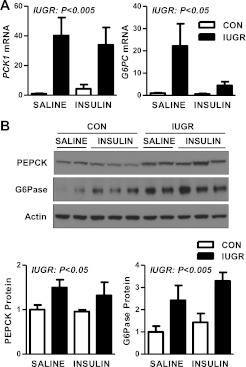
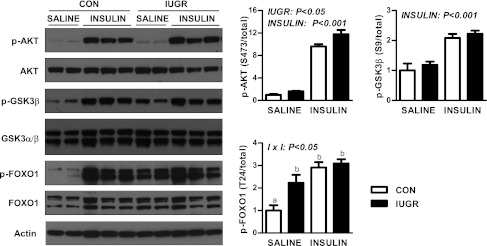
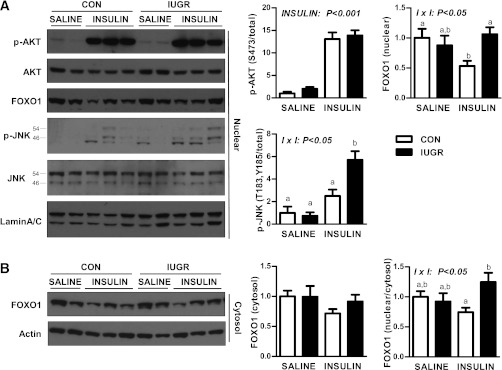
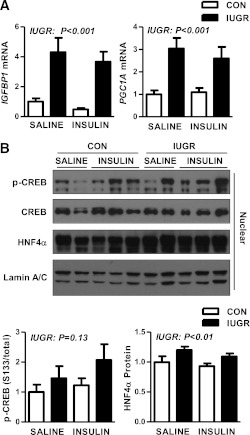
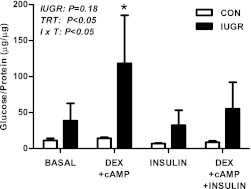
Intrauterine growth restriction (IUGR) increases the risk for metabolic disease and diabetes, although the developmental origins of this remain unclear. We measured glucose metabolism during basal and insulin clamp periods in a fetal sheep model of placental insufficiency and IUGR. Compared with control fetuses (CON), fetuses with IUGR had increased basal glucose production rates and hepatic PEPCK and glucose-6-phosphatase expression, which were not suppressed by insulin. In contrast, insulin significantly increased peripheral glucose utilization rates in CON and IUGR fetuses. Insulin robustly activated AKT, GSK3β, and forkhead box class O (FOXO)1 in CON and IUGR fetal livers. IUGR livers, however, had increased basal FOXO1 phosphorylation, nuclear FOXO1 expression, and Jun NH(2)-terminal kinase activation during hyperinsulinemia. Expression of peroxisome proliferator-activated receptor γ coactivator 1α and hepatocyte nuclear factor-4α were increased in IUGR livers during basal and insulin periods. Cortisol and norepinephrine concentrations were positively correlated with glucose production rates. Isolated IUGR hepatocytes maintained increased glucose production in culture. In summary, fetal sheep with IUGR have increased hepatic glucose production, which is not suppressed by insulin despite insulin sensitivity for peripheral glucose utilization. These data are consistent with a novel mechanism involving persistent transcriptional activation in the liver that seems to be unique in the fetus with IUGR.
Figures
References
-
- Symonds ME, Sebert SP, Hyatt MA, Budge H. Nutritional programming of the metabolic syndrome. Nat Rev Endocrinol 2009;5:604–610 - PubMed
-
- Martin-Gronert MS, Ozanne SE. Experimental IUGR and later diabetes. J Intern Med 2007;261:437–452 - PubMed
-
- Limesand SW, Rozance PJ, Zerbe GO, Hutton JC, Hay WW., Jr Attenuated insulin release and storage in fetal sheep pancreatic islets with intrauterine growth restriction. Endocrinology 2006;147:1488–1497 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
