

Direct control of brown adipose tissue thermogenesis by central nervous system glucagon-like peptide-1 receptor signaling
- PMID: 22933116
- PMCID: PMC3478556
- DOI: 10.2337/db11-1556
Direct control of brown adipose tissue thermogenesis by central nervous system glucagon-like peptide-1 receptor signaling
Abstract
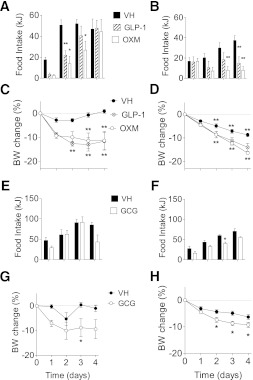
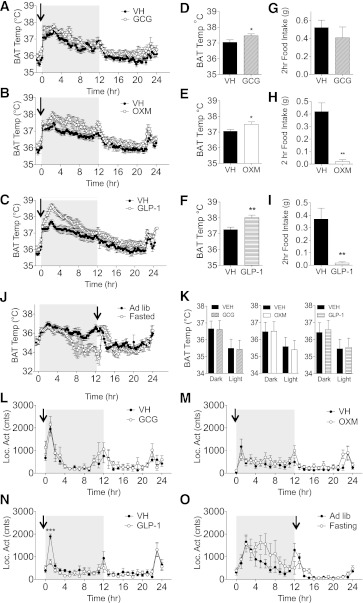
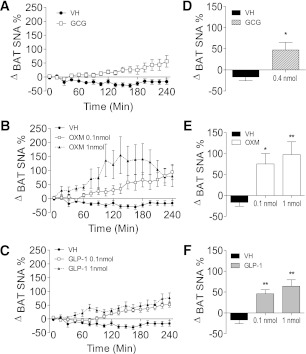
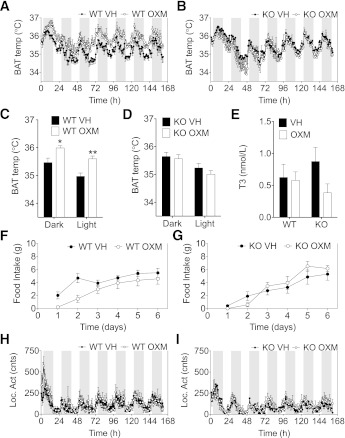
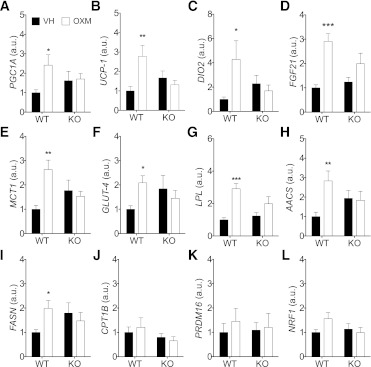
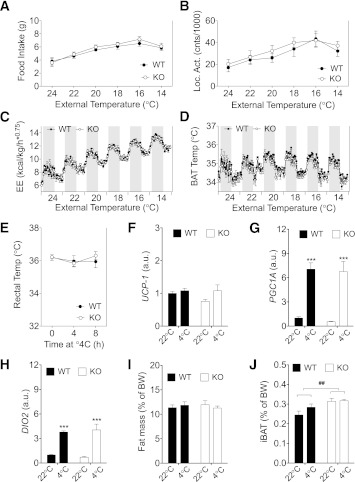
We studied interscapular brown adipose tissue (iBAT) activity in wild-type (WT) and glucagon-like peptide 1 receptor (GLP-1R)-deficient mice after the administration of the proglucagon-derived peptides (PGDPs) glucagon-like peptide (GLP-1), glucagon (GCG), and oxyntomodulin (OXM) directly into the brain. Intracerebroventricular injection of PGDPs reduces body weight and increases iBAT thermogenesis. This was independent of changes in feeding and insulin responsiveness but correlated with increased activity of sympathetic fibers innervating brown adipose tissue (BAT). Despite being a GCG receptor agonist, OXM requires GLP-1R activation to induce iBAT thermogenesis. The increase in thermogenesis in WT mice correlates with increased expression of genes upregulated by adrenergic signaling and required for iBAT thermogenesis, including PGC1a and UCP-1. In spite of the increase in iBAT thermogenesis induced by GLP-1R activation in WT mice, Glp1r(-/-) mice exhibit a normal response to cold exposure, demonstrating that endogenous GLP-1R signaling is not essential for appropriate thermogenic response after cold exposure. Our data suggest that the increase in BAT thermogenesis may be an additional mechanism whereby pharmacological GLP-1R activation controls energy balance.
Figures
References
-
- Wren AM, Bloom SR. Gut hormones and appetite control. Gastroenterology 2007;132:2116–2130 - PubMed
-
- Astrup A, Rössner S, Van Gaal L, et al. NN8022-1807 Study Group Effects of liraglutide in the treatment of obesity: a randomised, double-blind, placebo-controlled study. Lancet 2009;374:1606–1616 - PubMed
-
- Heppner KM, Habegger KM, Day J, et al. Glucagon regulation of energy metabolism. Physiol Behav 2010;100:545–548 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
