

IRF4 promotes cutaneous dendritic cell migration to lymph nodes during homeostasis and inflammation
- PMID: 22933627
- PMCID: PMC3448873
- DOI: 10.4049/jimmunol.1102613
IRF4 promotes cutaneous dendritic cell migration to lymph nodes during homeostasis and inflammation
Abstract
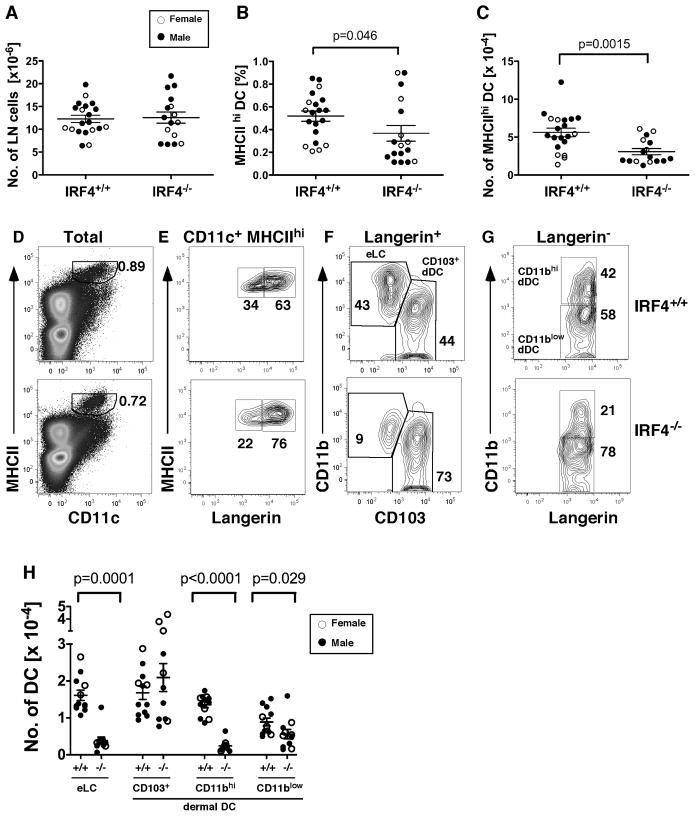
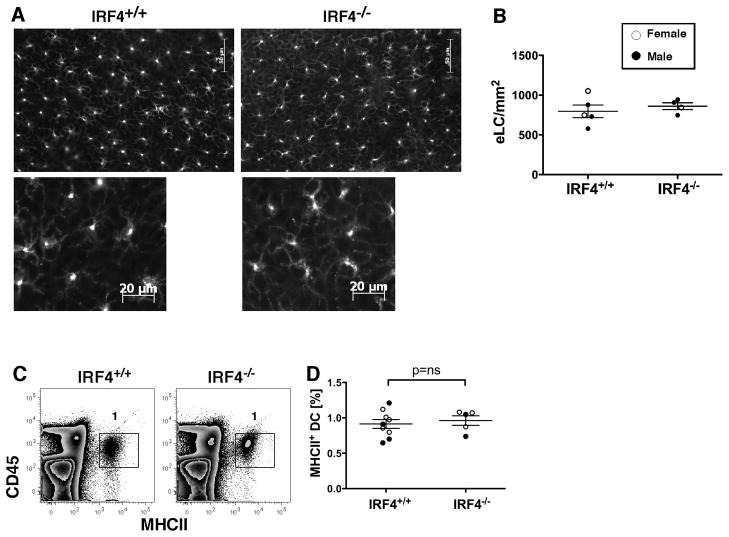
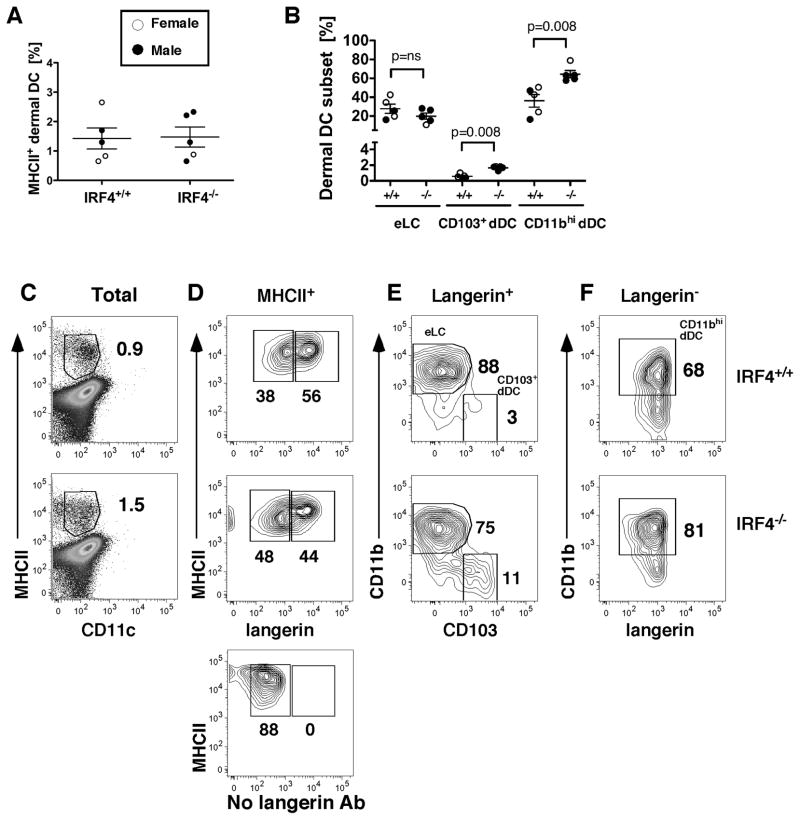
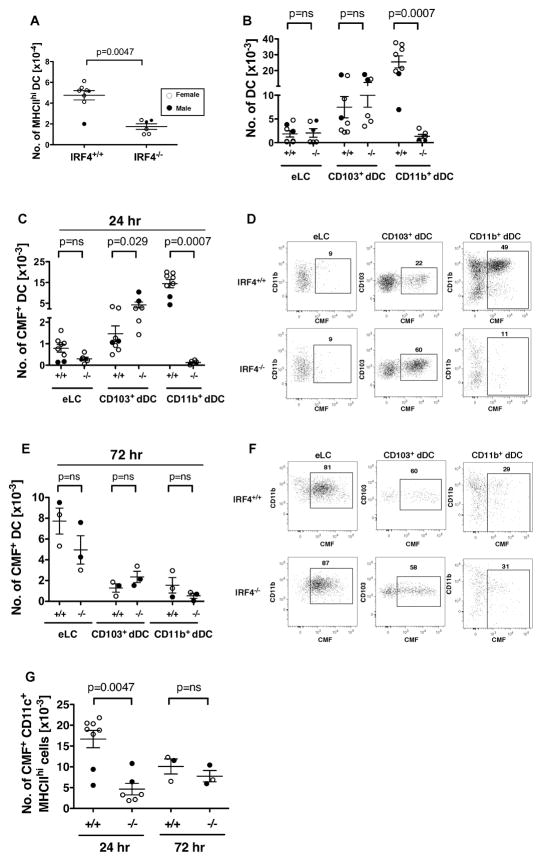
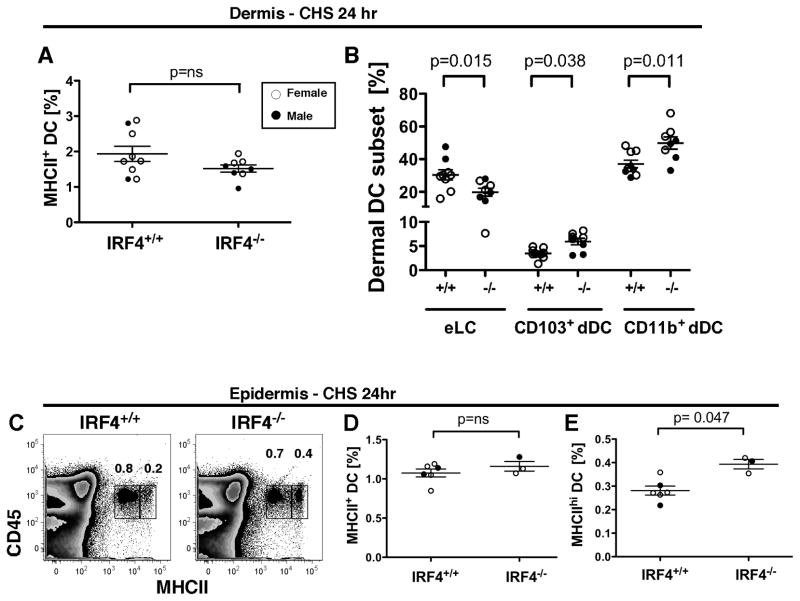
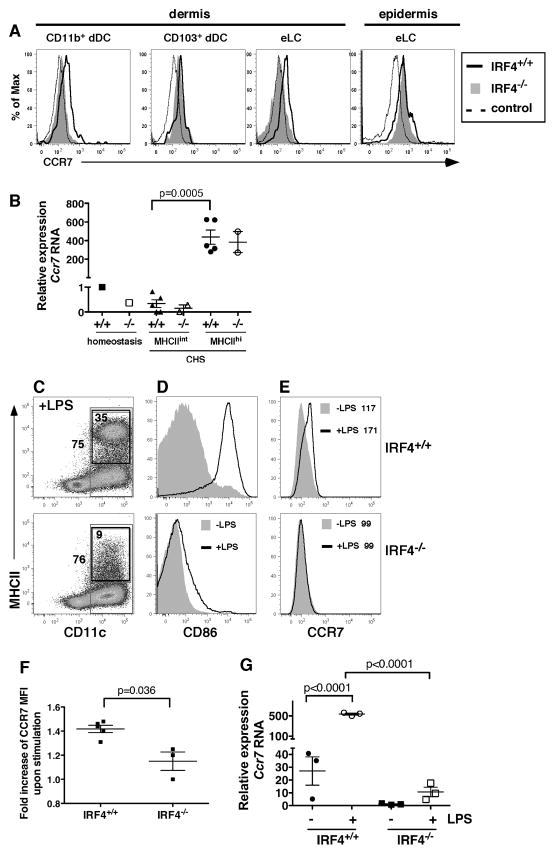
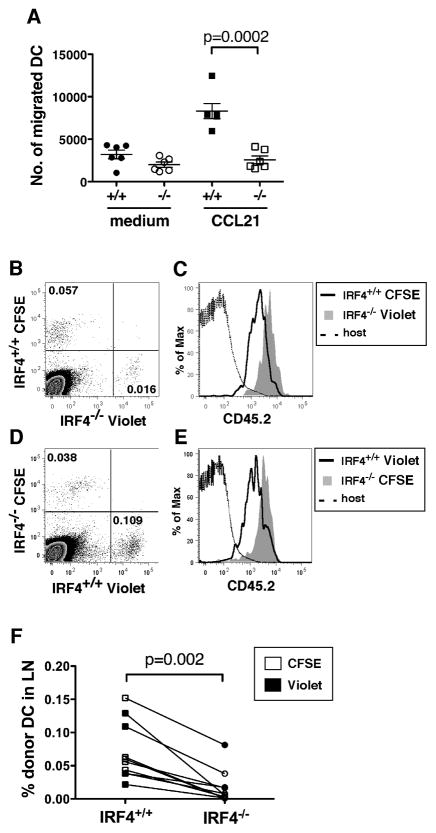
Migration of resident dendritic cells (DC) from the skin to local lymph nodes (LN) triggers T cell-mediated immune responses during cutaneous infection, autoimmune disease, and vaccination. In this study, we investigated whether the development and migration of skin-resident DC were regulated by IFN regulatory factor 4 (IRF4), a transcription factor that is required for the development of CD11b(+) splenic DC. We found that the skin of naive IRF4(-/-) mice contained normal numbers of epidermal Langerhans cells (eLC) and increased numbers of CD11b(+) and CD103(+) dermal DC (dDC) populations, indicating that tissue DC development and skin residency is not disrupted by IRF4 deficiency. In contrast, numbers of migratory eLC and CD11b(+) dDC were significantly reduced in the cutaneous LN of IRF4(-/-) mice, suggesting a defect in constitutive migration from the dermis during homeostasis. Upon induction of skin inflammation, CD11b(+) dDC in IRF4(-/-) mice did not express the chemokine receptor CCR7 and failed to migrate to cutaneous LN, whereas the migration of eLC was only mildly impaired. Thus, although dispensable for their development, IRF4 is crucial for the CCR7-mediated migration of CD11b(+) dDC, a predominant population in murine and human skin that plays a vital role in normal and pathogenic cutaneous immunity.
Conflict of interest statement
None.
Figures
References
-
- Ohl L, Mohaupt M, Czeloth N, Hintzen G, Kiafard Z, Zwirner J, Blankenstein T, Henning G, Forster R. CCR7 governs skin dendritic cell migration under inflammatory and steady-state conditions. Immunity. 2004;21:279–288. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
