

Virus-host swinging party in the oceans: Incorporating biological complexity into paradigms of antagonistic coexistence
- PMID: 22934242
- PMCID: PMC3429526
- DOI: 10.4161/mge.20031
Virus-host swinging party in the oceans: Incorporating biological complexity into paradigms of antagonistic coexistence
Abstract
Bacteria and their viruses (phages) are antagonists, yet have coexisted in nature for billions of years. Models proposed to explain the paradox of antagonistic coexistence generally reach two types of solutions: Arms race-like dynamics that lead to hosts and viruses with increasing resistance and infection ranges; and population fluctuations between diverse host and viral types due to a metabolic cost of resistance. Recently, we found that populations of the marine cyanobacterium, Prochlorococcus, consist of cells with extreme hypervariability in gene sequence and gene content in a viral susceptibility region of the genome. Furthermore, we found a novel cost of resistance where resistance to one set of viruses is accompanied by changes in infection dynamics by other viruses. In this combined mini-review and commentary paper we discuss these findings in the context of existing ecological, evolutionary and genetic models of host-virus coexistence. We suggest that this coexistence is governed mainly by fluctuations between microbial subpopulations with differing viral susceptibility regions and that these fluctuations are driven by both metabolic and enhanced infection costs of resistance. Furthermore, we suggest that enhanced infection leads to passive host-switching by viruses, preventing the development of hosts with universal resistance. These findings highlight the vital importance of community complexity for host-virus coexistence.
Figures
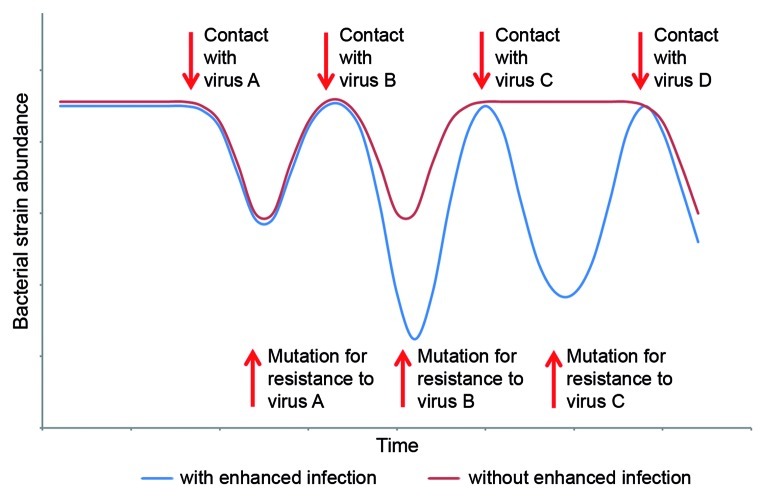
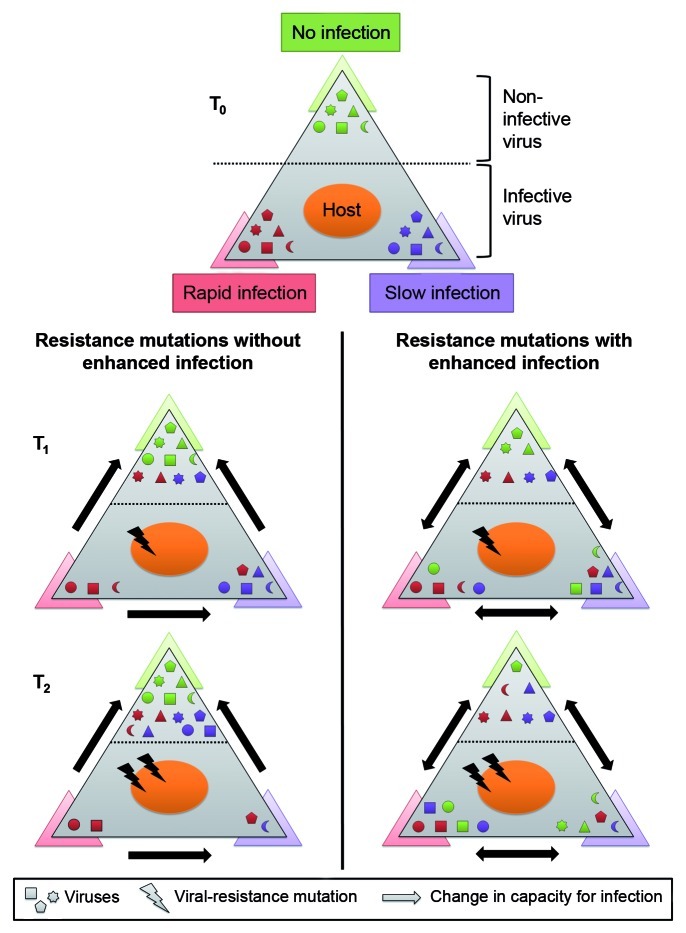
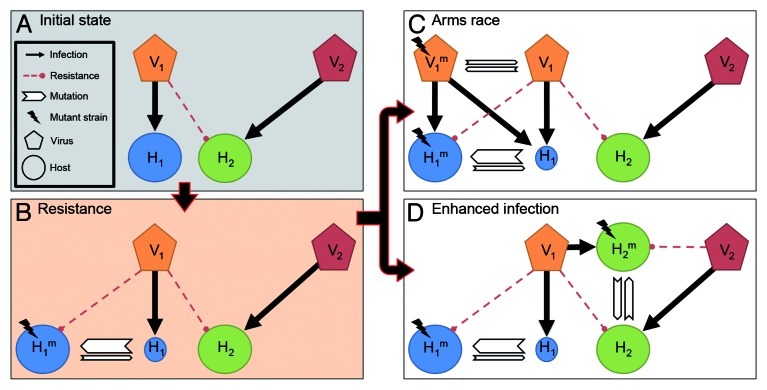
Comment on
- Avrani S, Wurtzel O, Sharon I, Sorek R, Lindell D. Genomic island variability facilitates Prochlorococcus-virus coexistence. Nature. 2011;474:604–8. doi: 10.1038/nature10172.
References
-
- Thompson JN. The Coevolutionary Process. Chicago, IL: University of Chicago Press, 1994:253-75.
-
- Bohannan BJM, Lenski RE. Linking genetic change to community evolution: insights from studies of bacteria and bacteriophage. Ecol Lett. 2000;3:362–77. doi: 10.1046/j.1461-0248.2000.00161.x. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources