

A putative role for amino acid permeases in sink-source communication of barley tissues uncovered by RNA-seq
- PMID: 22935196
- PMCID: PMC3495740
- DOI: 10.1186/1471-2229-12-154
A putative role for amino acid permeases in sink-source communication of barley tissues uncovered by RNA-seq
Abstract
Background: The majority of nitrogen accumulating in cereal grains originates from proteins remobilised from vegetative organs. However, interactions between grain filling and remobilisation are poorly understood. We used transcriptome large-scale pyrosequencing of flag leaves, glumes and developing grains to identify cysteine peptidase and N transporter genes playing a role in remobilisation and accumulation of nitrogen in barley.
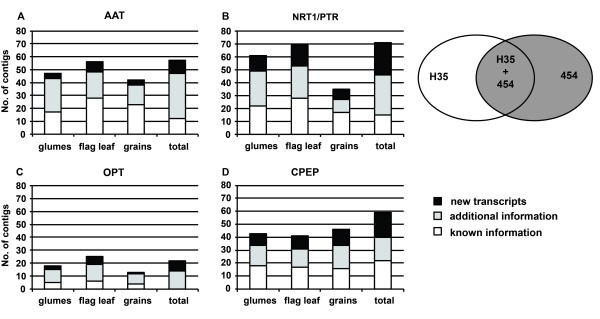
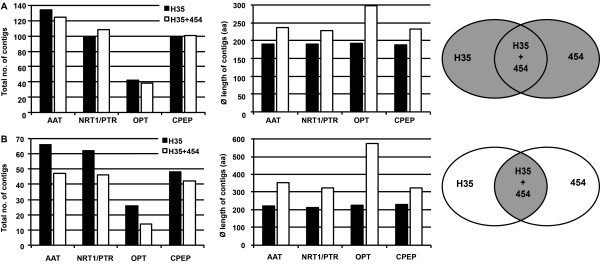
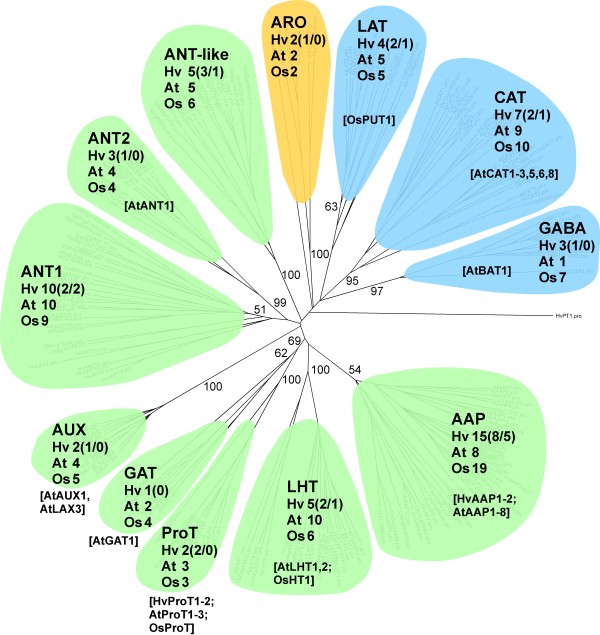
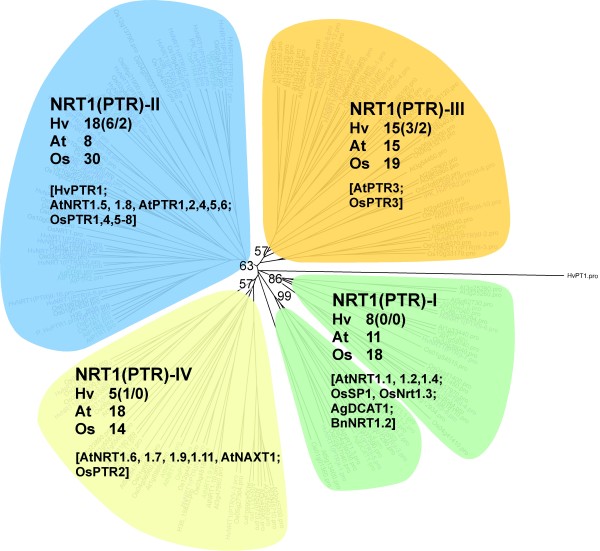
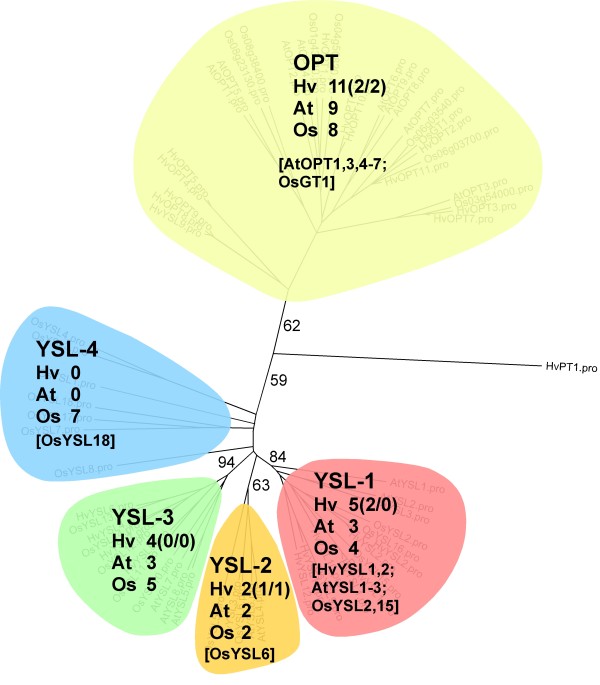
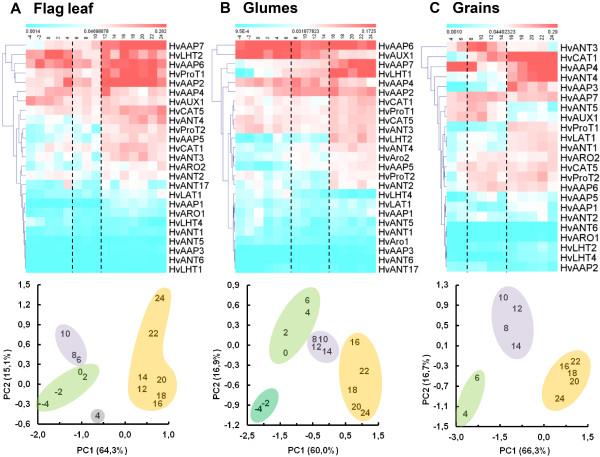
Results: Combination of already known and newly derived sequence information reduced redundancy, increased contig length and identified new members of cysteine peptidase and N transporter gene families. The dataset for N transporter genes was aligned with N transporter amino acid sequences of rice and Arabidopsis derived from Aramemnon database. 57 AAT, 45 NRT1/PTR and 22 OPT unigenes identified by this approach cluster to defined subgroups in the respective phylogenetic trees, among them 25 AAT, 8 NRT1/PTR and 5 OPT full-length sequences. Besides, 59 unigenes encoding cysteine peptidases were identified and subdivided into different families of the papain cysteine peptidase clade. Expression profiling of full-length AAT genes highlighted amino acid permeases as the group showing highest transcriptional activity. HvAAP2 and HvAAP6 are highly expressed in vegetative organs whereas HvAAP3 is grain-specific. Sequence similarities cluster HvAAP2 and the putative transporter HvAAP6 together with Arabidopsis transporters, which are involved in long-distance transfer of amino acids. HvAAP3 is closely related to AtAAP1 and AtAAP8 playing a role in supplying N to developing seeds. An important role in amino acid re-translocation can be considered for HvLHT1 and HvLHT2 which are specifically expressed in glumes and flag leaves, respectively. PCA and K-means clustering of AAT transcript data revealed coordinate developmental stages in flag leaves, glumes and grains. Phloem-specific metabolic compounds are proposed that might signal high grain demands for N to distantly located plant organs.
Conclusions: The approach identified cysteine peptidases and specific N transporters of the AAT family as obviously relevant for grain filling and thus, grain yield and quality in barley. Up to now, information is based only on transcript data. To make it relevant for application, the role of identified candidates in sink-source communication has to be analysed in more detail.
Figures

References
-
- Peoples MBDM. The interlplay between proteolysis and amino acid metabolism during senescence and aging in plants. New York: AcademicPress Inc; 1988.
-
- Peeters KMU, Van Laere AJ. Amino acid metabolism associated with N-mobilization from the flag leaf of wheat (Triticum aestivum L.) during grain development. Plant Cell Environ. 1994;17(2):131–141. doi: 10.1111/j.1365-3040.1994.tb00276.x. - DOI
-
- Masclaux-Daubresse C, Reisdorf-Cren M, Orsel M. Leaf nitrogen remobilisation for plant development and grain filling. Plant Biology. 2008;10:23–36. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
