

Noncanonical control of C. elegans germline apoptosis by the insulin/IGF-1 and Ras/MAPK signaling pathways
- PMID: 22935616
- PMCID: PMC3524627
- DOI: 10.1038/cdd.2012.101
Noncanonical control of C. elegans germline apoptosis by the insulin/IGF-1 and Ras/MAPK signaling pathways
Abstract
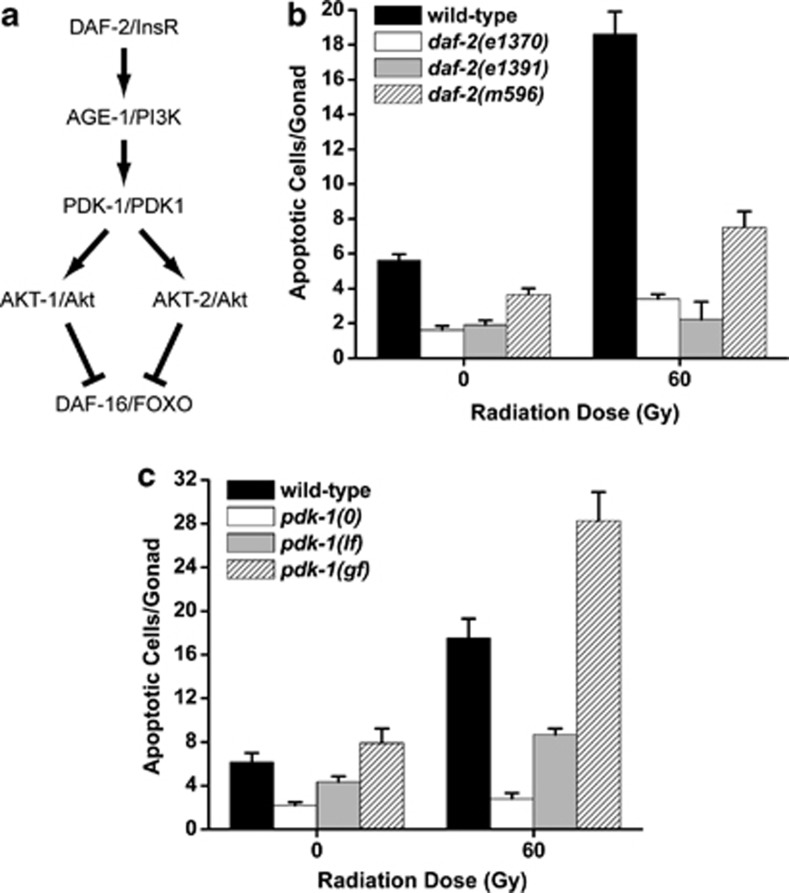
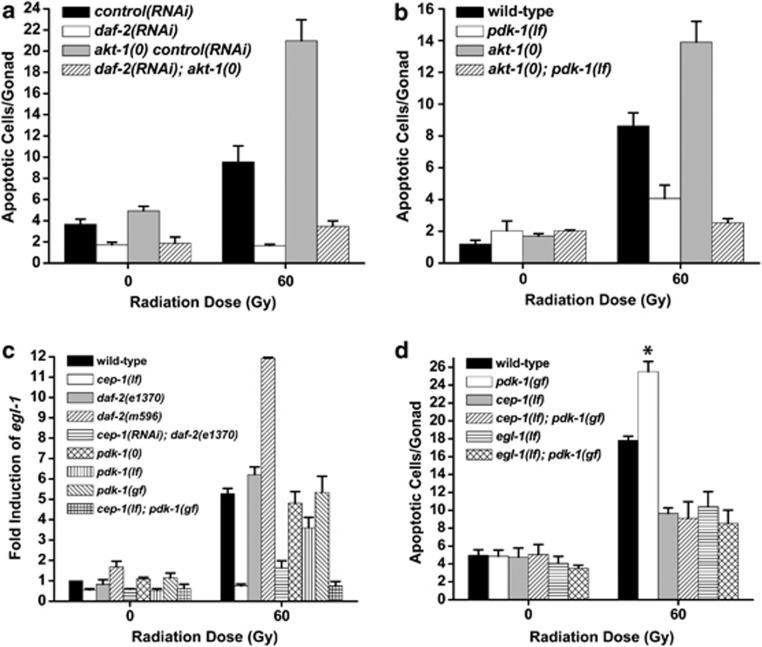
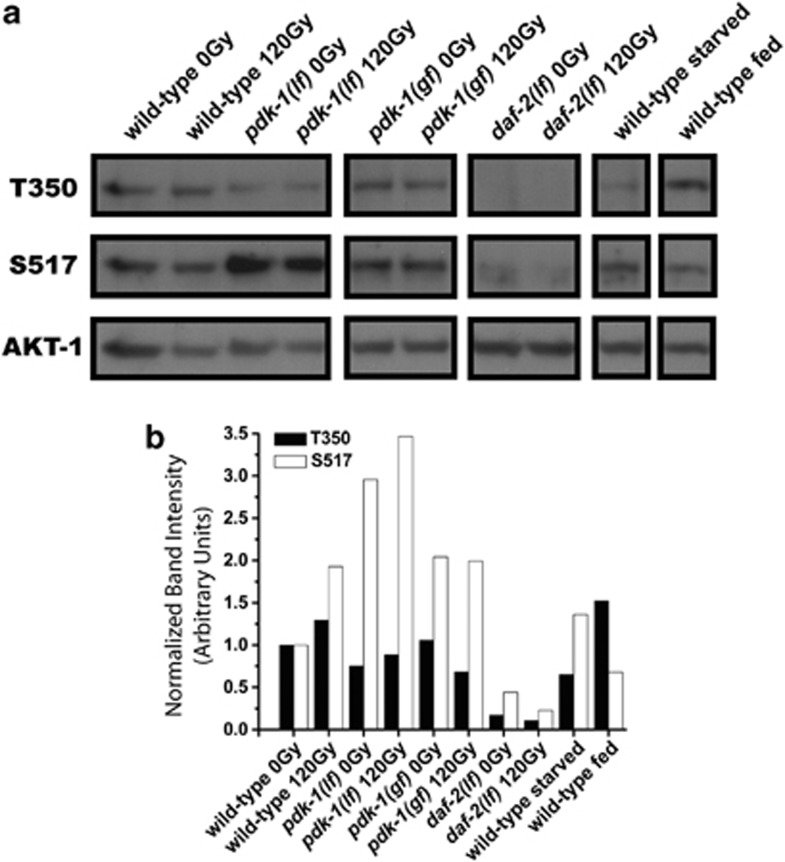
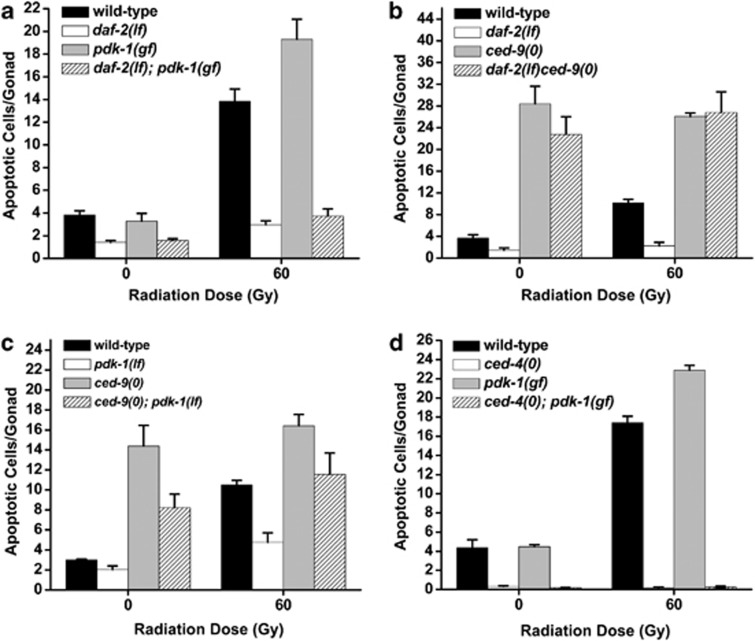
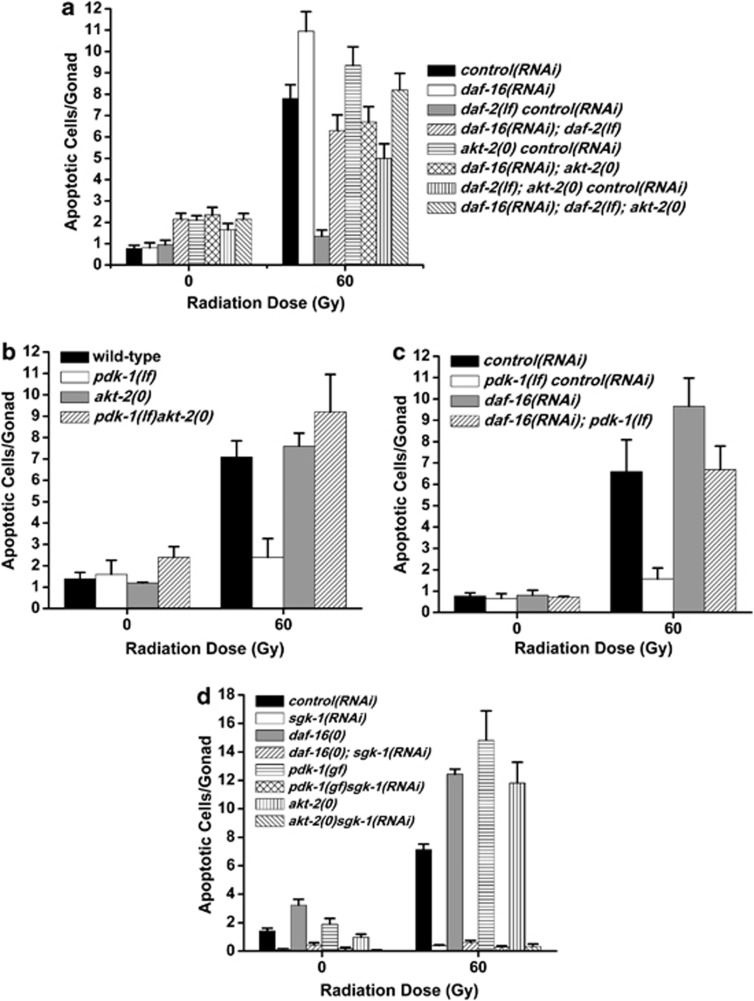
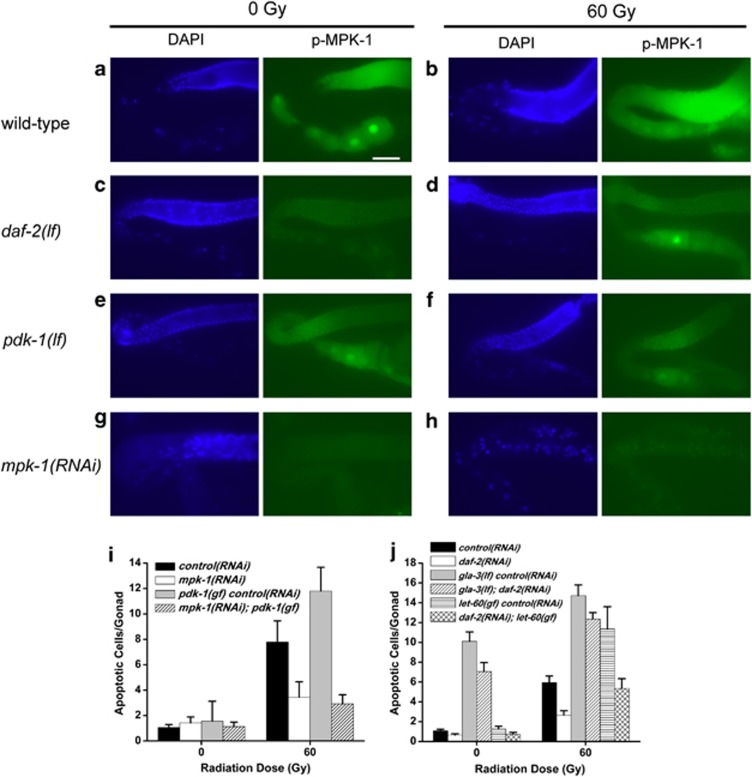
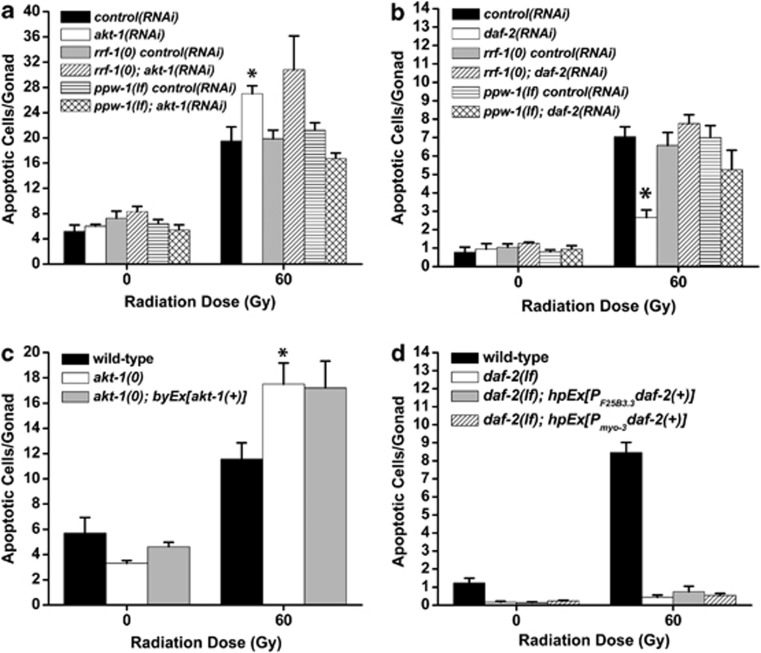
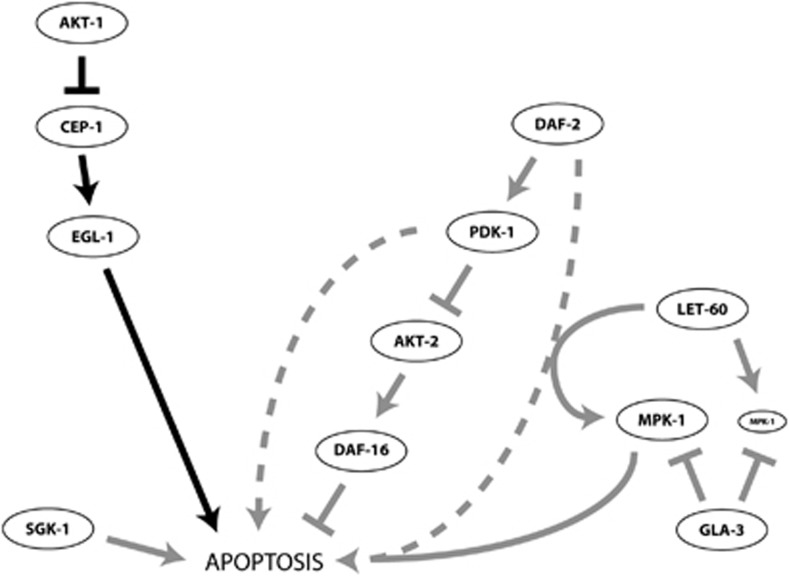
The insulin/IGF-1 pathway controls a number of physiological processes in the nematode worm Caenorhabditis elegans, including development, aging and stress response. We previously found that the Akt/PKB ortholog AKT-1 dampens the apoptotic response to genotoxic stress in the germline by negatively regulating the p53-like transcription factor CEP-1. Here, we report unexpected rearrangements to the insulin/IGF-1 pathway, whereby the insulin-like receptor DAF-2 and 3-phosphoinositide-dependent protein kinase PDK-1 oppose AKT-1 to promote DNA damage-induced apoptosis. While DNA damage does not affect phosphorylation at the PDK-1 site Thr350/Thr308 of AKT-1, it increased phosphorylation at Ser517/Ser473. Although ablation of daf-2 or pdk-1 completely suppressed akt-1-dependent apoptosis, the transcriptional activation of CEP-1 was unaffected, suggesting that daf-2 and pdk-1 act independently or downstream of cep-1 and akt-1. Ablation of the akt-1 paralog akt-2 or the downstream target of the insulin/IGF-1 pathway daf-16 (a FOXO transcription factor) restored sensitivity to damage-induced apoptosis in daf-2 and pdk-1 mutants. In addition, daf-2 and pdk-1 mutants have reduced levels of phospho-MPK-1/ERK in their germ cells, indicating that the insulin/IGF-1 pathway promotes Ras signaling in the germline. Ablation of the Ras effector gla-3, a negative regulator of mpk-1, restored sensitivity to apoptosis in daf-2 mutants, suggesting that gla-3 acts downstream of daf-2. In addition, the hypersensitivity of let-60/Ras gain-of-function mutants to damage-induced apoptosis was suppressed to wild-type levels by ablation of daf-2. Thus, insulin/IGF-1 signaling selectively engages AKT-2/DAF-16 to promote DNA damage-induced germ cell apoptosis downstream of CEP-1 through the Ras pathway.
Figures
References
-
- Vivanco I, Sawyers CL. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev Cancer. 2002;2:489–501. - PubMed
-
- Contessa JN, Hampton J, Lammering G, Mikkelsen RB, Dent P, Valerie K, et al. Ionizing radiation activates Erb-B receptor dependent Akt and p70 S6 kinase signaling in carcinoma cells. Oncogene. 2002;21:4032–4041. - PubMed
-
- Brognard J, Clark AS, Ni Y, Dennis PA. Akt/protein kinase B is constitutively active in non-small cell lung cancer cells and promotes cellular survival and resistance to chemotherapy and radiation. Cancer Res. 2001;61:3986–3997. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
