

Waxy phenotype evolution in the allotetraploid cereal broomcorn millet: mutations at the GBSSI locus in their functional and phylogenetic context
- PMID: 22936718
- PMCID: PMC3533377
- DOI: 10.1093/molbev/mss209
Waxy phenotype evolution in the allotetraploid cereal broomcorn millet: mutations at the GBSSI locus in their functional and phylogenetic context
Abstract
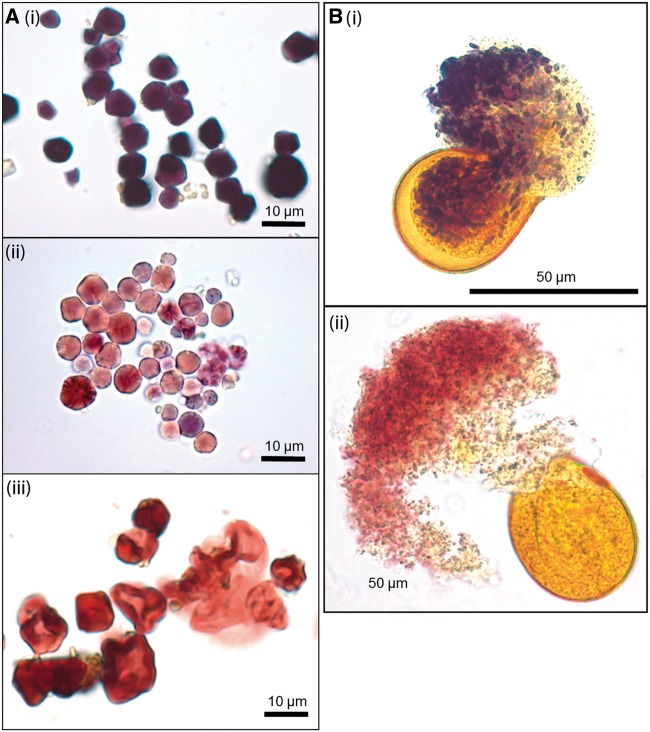
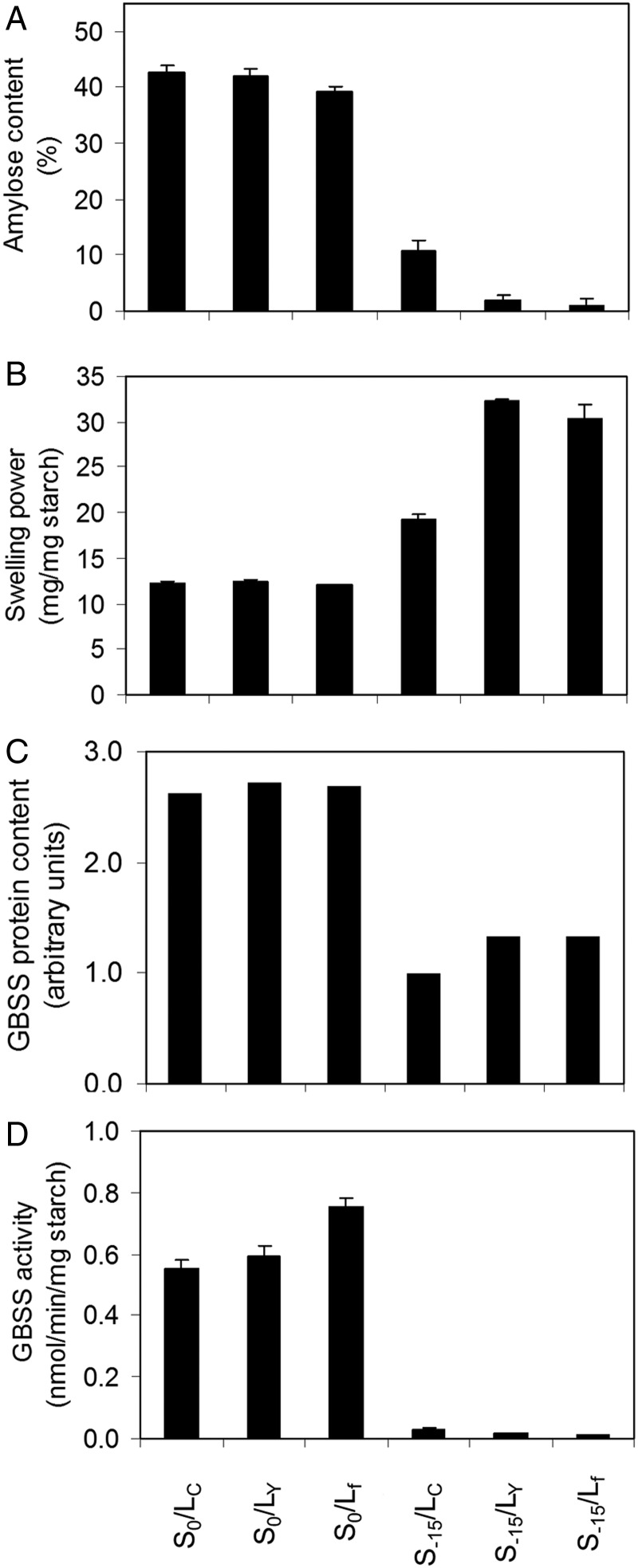
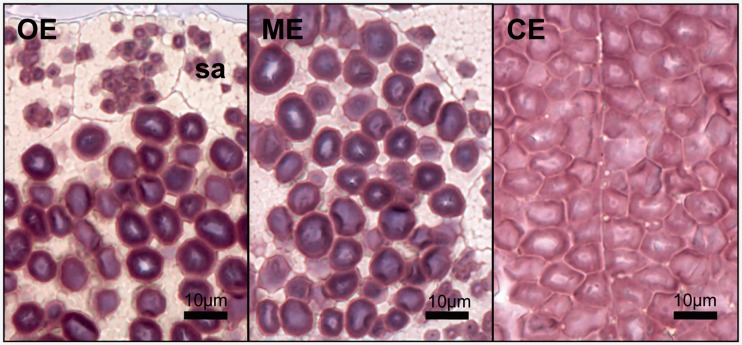
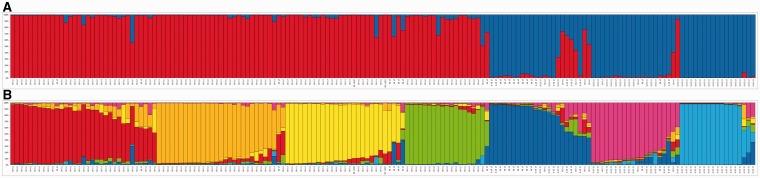
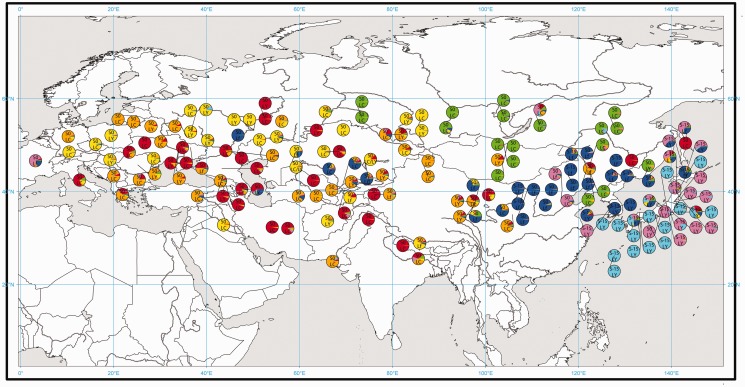
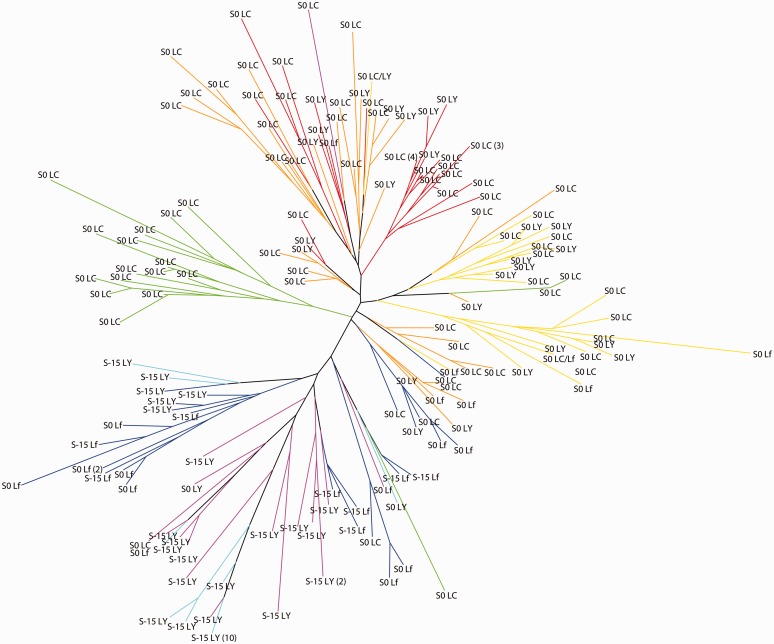
Waxy mutants, in which endosperm starch contains ~100% amylopectin rather than the wild-type composition of ~70% amylopectin and ~30% amylose, occur in many domesticated cereals. The cultivation of waxy varieties is concentrated in east Asia, where there is a culinary preference for glutinous-textured foods that may have developed from ancient food processing traditions. The waxy phenotype results from mutations in the GBSSI gene, which catalyzes amylose synthesis. Broomcorn or proso millet (Panicum miliaceum L.) is one of the world's oldest cultivated cereals, which spread across Eurasia early in prehistory. Recent phylogeographic analysis has shown strong genetic structuring that likely reflects ancient expansion patterns. Broomcorn millet is highly unusual in being an allotetraploid cereal with fully waxy varieties. Previous work characterized two homeologous GBSSI loci, with multiple alleles at each, but could not determine whether both loci contributed to GBSSI function. We first tested the relative contribution of the two GBSSI loci to amylose synthesis and second tested the association between GBSSI alleles and phylogeographic structure inferred from simple sequence repeats (SSRs). We evaluated the phenotype of all known GBSSI genotypes in broomcorn millet by assaying starch composition and protein function. The results showed that the GBSSI-S locus is the major locus controlling endosperm amylose content, and the GBSSI-L locus has strongly reduced synthesis capacity. We genotyped 178 individuals from landraces from across Eurasia for the 2 GBSSI and 16 SSR loci and analyzed phylogeographic structuring and the geographic and phylogenetic distribution of GBSSI alleles. We found that GBSSI alleles have distinct spatial distributions and strong associations with particular genetic clusters defined by SSRs. The combination of alleles that results in a partially waxy phenotype does not exist in landrace populations. Our data suggest that broomcorn millet is a system in the process of becoming diploidized for the GBSSI locus responsible for grain amylose. Mutant alleles show some exchange between genetic groups, which was favored by selection for the waxy phenotype in particular regions. Partially waxy phenotypes were probably selected against-this unexpected finding shows that better understanding is needed of the human biology of this phenomenon that distinguishes cereal use in eastern and western cultures.
Figures
Similar articles
-
Molecular basis of the waxy endosperm starch phenotype in broomcorn millet (Panicum miliaceum L.).Mol Biol Evol. 2010 Jul;27(7):1478-94. doi: 10.1093/molbev/msq040. Epub 2010 Feb 5. Mol Biol Evol. 2010. PMID: 20139147 Free PMC article.
-
Transcriptome analysis reveals new insights in the starch biosynthesis of non-waxy and waxy broomcorn millet (Panicum miliaceum L.).Int J Biol Macromol. 2023 Mar 1;230:123155. doi: 10.1016/j.ijbiomac.2023.123155. Epub 2023 Jan 4. Int J Biol Macromol. 2023. PMID: 36610580
-
Molecular insights on the origin and development of waxy genotypes in major crop plants.Brief Funct Genomics. 2024 May 15;23(3):193-213. doi: 10.1093/bfgp/elad035. Brief Funct Genomics. 2024. PMID: 38751352 Review.
-
Waxy allele diversification in foxtail millet (Setaria italica) landraces of Taiwan.PLoS One. 2018 Dec 31;13(12):e0210025. doi: 10.1371/journal.pone.0210025. eCollection 2018. PLoS One. 2018. PMID: 30596758 Free PMC article.
-
Proso-millet starch: Properties, functionality, and applications.Int J Biol Macromol. 2021 Nov 1;190:960-968. doi: 10.1016/j.ijbiomac.2021.09.064. Epub 2021 Sep 16. Int J Biol Macromol. 2021. PMID: 34536472 Review.
Cited by
-
Harnessing haploid-inducer mediated genome editing for accelerated maize variety development.Plant Biotechnol J. 2025 May;23(5):1604-1614. doi: 10.1111/pbi.14608. Epub 2025 Feb 12. Plant Biotechnol J. 2025. PMID: 39936495 Free PMC article.
-
Expediting Next-Generation Hybrid Technology in Recalcitrant Maize Inbreds through In Vivo Targeted Activity of CRISPR/Cas9.Int J Mol Sci. 2024 May 27;25(11):5832. doi: 10.3390/ijms25115832. Int J Mol Sci. 2024. PMID: 38892019 Free PMC article.
-
Crop plants as models for understanding plant adaptation and diversification.Front Plant Sci. 2013 Aug 1;4:290. doi: 10.3389/fpls.2013.00290. eCollection 2013. Front Plant Sci. 2013. PMID: 23914199 Free PMC article.
-
Generation of new rice germplasms with low amylose content by CRISPR/CAS9-targeted mutagenesis of the FLOURY ENDOSPERM 2 gene.Front Plant Sci. 2023 Mar 13;14:1138523. doi: 10.3389/fpls.2023.1138523. eCollection 2023. Front Plant Sci. 2023. PMID: 36993856 Free PMC article.
-
Parallelism and convergence in post-domestication adaptation in cereal grasses.Philos Trans R Soc Lond B Biol Sci. 2019 Jul 22;374(1777):20180245. doi: 10.1098/rstb.2018.0245. Epub 2019 Jun 3. Philos Trans R Soc Lond B Biol Sci. 2019. PMID: 31154975 Free PMC article. Review.
References
-
- Åkerberg A, Liljeberg H, Björck I. Effects of amylose/amylopectin ratio and baking conditions on resistant starch formation and glycaemic indices. J Cereal Sci. 1998;28:71–80.
-
- Araki M, Numaoka A, Kawase M, Fukunaga K. Origin of waxy common millet, Panicum miliaceum L. in Japan. Genet Resour Crop Evol. Forthcoming doi 10.1007/s10722-011-9755-9.
-
- Baltensperger DD. Progress with proso, pearl and other millets. In: Janick J, Whipkey A, editors. Trends in new crops and new uses. Alexandria (VA): ASHS Press; 2002. pp. 100–103.
-
- Boutin-Ganache I, Raposo M, Raymond M, Deschepper CF. M13-tailed primers improve the readability and usability of microsatellite analyses performed with two different allele-sizing methods. Biotechniques. 2001;31:24–28. - PubMed
-
- Boyer CD, Shannon JC, Garwood DL, Creech RG. Changes in starch granule size and amylose percentage during kernel development in several Zea-Mays-L genotypes. Cereal Chem. 1976;53:327–337.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
