

Comparative structure and biomechanics of plant primary and secondary cell walls
- PMID: 22936943
- PMCID: PMC3424969
- DOI: 10.3389/fpls.2012.00204
Comparative structure and biomechanics of plant primary and secondary cell walls
Abstract
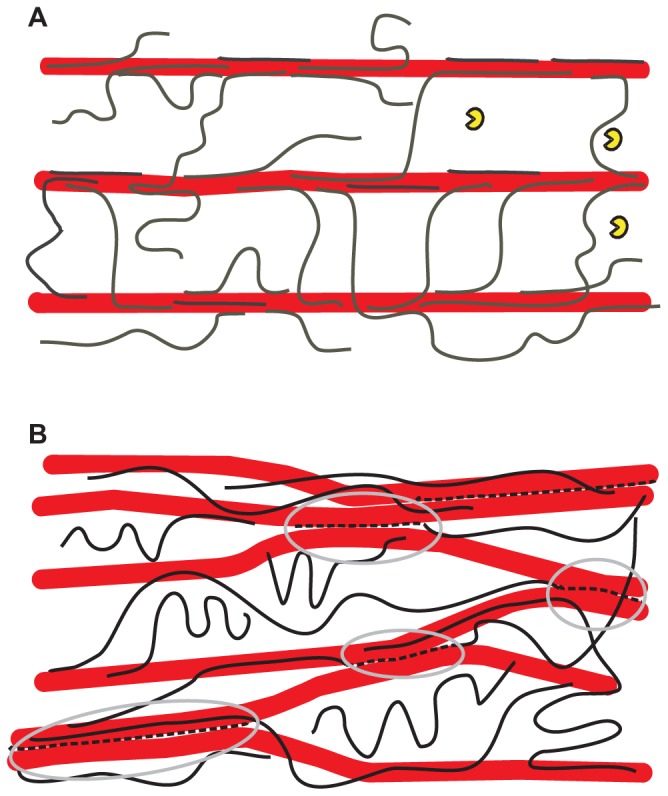
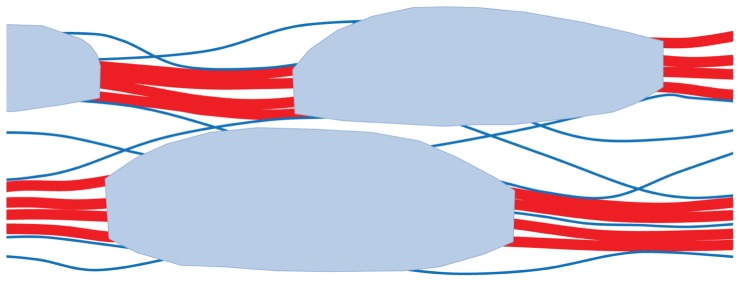
Recent insights into the physical biology of plant cell walls are reviewed, summarizing the essential differences between primary and secondary cell walls and identifying crucial gaps in our knowledge of their structure and biomechanics. Unexpected parallels are identified between the mechanism of expansion of primary cell walls during growth and the mechanisms by which hydrated wood deforms under external tension. There is a particular need to revise current "cartoons" of plant cell walls to be more consistent with data from diverse approaches and to go beyond summarizing limited aspects of cell walls, serving instead as guides for future experiments and for the application of new techniques.
Keywords: cellulose; creep; deformation; growth; primary cell walls; secondary cell walls; xyloglucan.
Figures
References
-
- Abasolo W., Eder M., Yamauchi K., Obel N., Reinecke A., Neumetzler L., Dunlop J. W., Mouille G., Pauly M., Hofte H., Burgert I. (2009). Pectin may hinder the unfolding of xyloglucan chains during cell deformation: implications of the mechanical performance of Arabidopsis hypocotyls with pectin alterations. Mol. Plant 2 990–999 - PubMed
-
- Altaner C. M., Jarvis M. C. (2008). Modelling polymer interactions of the “molecular Velcro” type in wood under mechanical stress. J. Theor. Biol. 253 434–445 - PubMed
-
- Assor C., Placet V., Chabbert B., Habrant A., Lapierre C., Pollet B., Perre P. (2009). Concomitant changes in viscoelastic properties and amorphous polymers during the hydrothermal treatment of hardwood and softwood. J. Agric. Food Chem. 57 6830–6837 - PubMed
-
- Atalla R. H., Hackney J. M., Uhlin I., Thompson N. S. (1993). Hemicelluloses as structure regulators in the aggregation of native cellulose. Int. J. Biol. Macromol. 15 109–112 - PubMed
LinkOut - more resources
Full Text Sources